

Neurite elongation is highly correlated with bulk forward translocation of microtubules
- PMID: 28779177
- PMCID: PMC5544698
- DOI: 10.1038/s41598-017-07402-6
Neurite elongation is highly correlated with bulk forward translocation of microtubules
Abstract
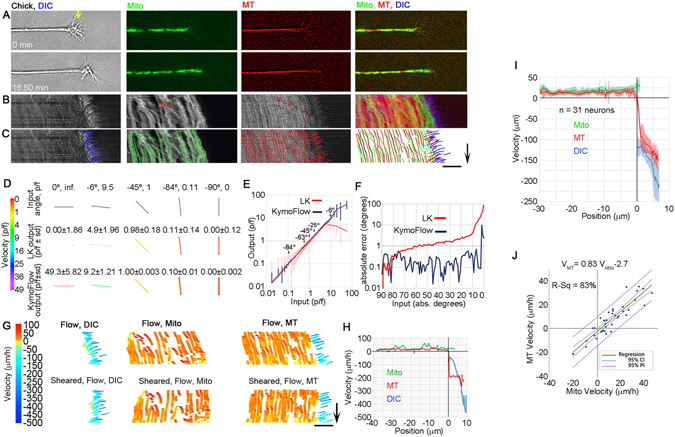
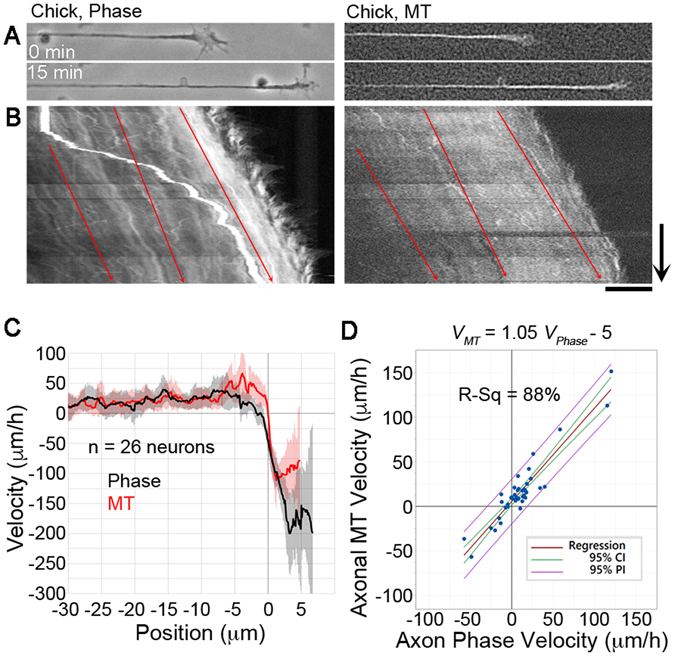
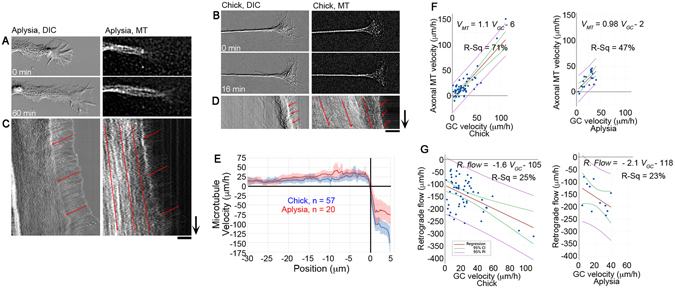
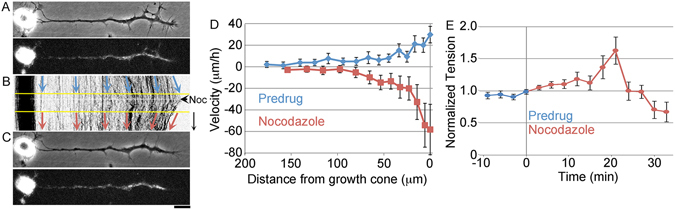
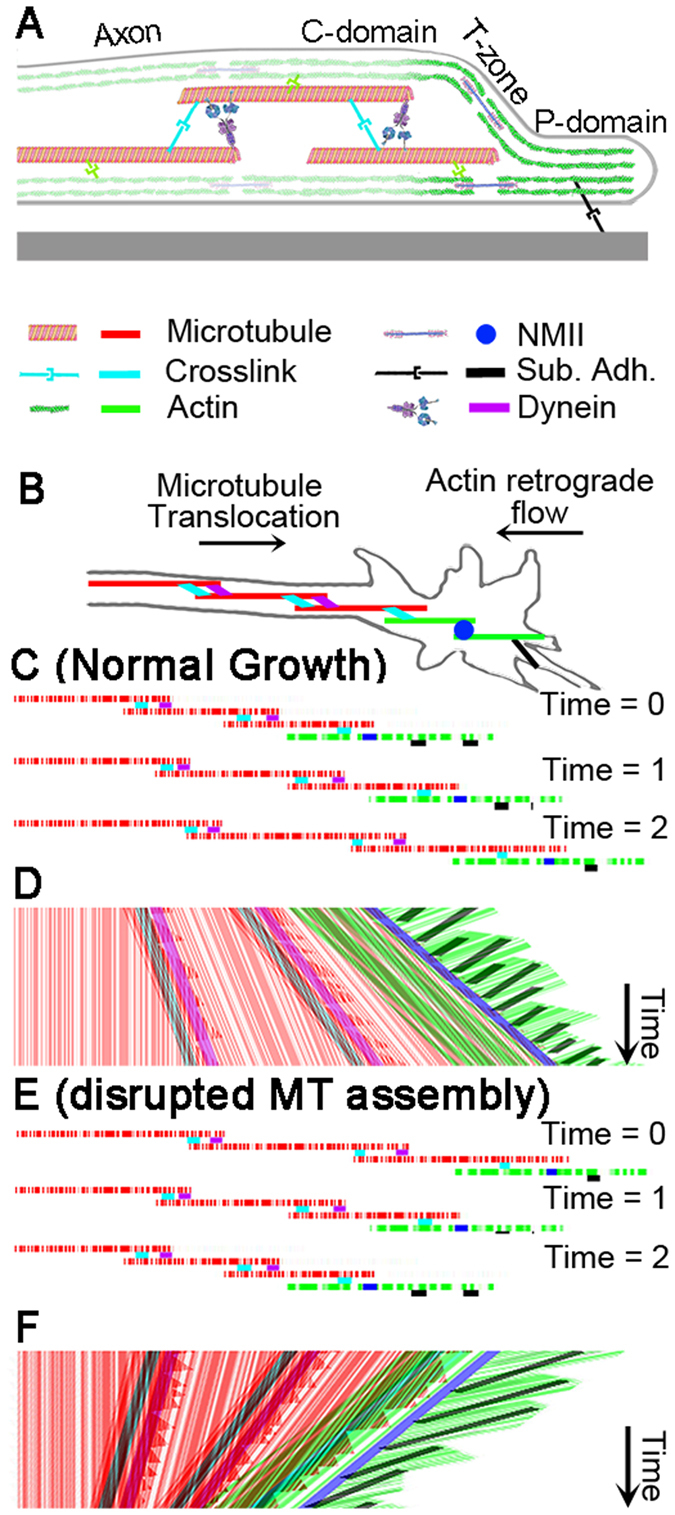
During the development of the nervous system and regeneration following injury, microtubules (MTs) are required for neurite elongation. Whether this elongation occurs primarily through tubulin assembly at the tip of the axon, the transport of individual MTs, or because MTs translocate forward in bulk is unclear. Using fluorescent speckle microscopy (FSM), differential interference contrast (DIC), and phase contrast microscopy, we tracked the movement of MTs, phase dense material, and docked mitochondria in chick sensory and Aplysia bag cell neurons growing rapidly on physiological substrates. In all cases, we find that MTs and other neuritic components move forward in bulk at a rate that on average matches the velocity of neurite elongation. To better understand whether and why MT assembly is required for bulk translocation, we disrupted it with nocodazole. We found this blocked the forward bulk advance of material along the neurite and was paired with a transient increase in axonal tension. This indicates that disruption of MT dynamics interferes with neurite outgrowth, not by disrupting the net assembly of MTs at the growth cone, but rather because it alters the balance of forces that power the bulk forward translocation of MTs.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
