

Evolution of complex adaptations in molecular systems
- PMID: 28782044
- PMCID: PMC5540182
- DOI: 10.1038/s41559-017-0228-1
Evolution of complex adaptations in molecular systems
Abstract
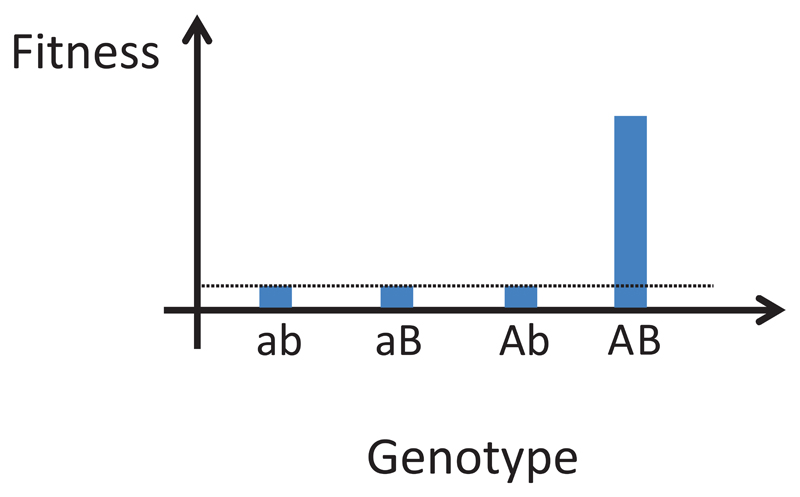
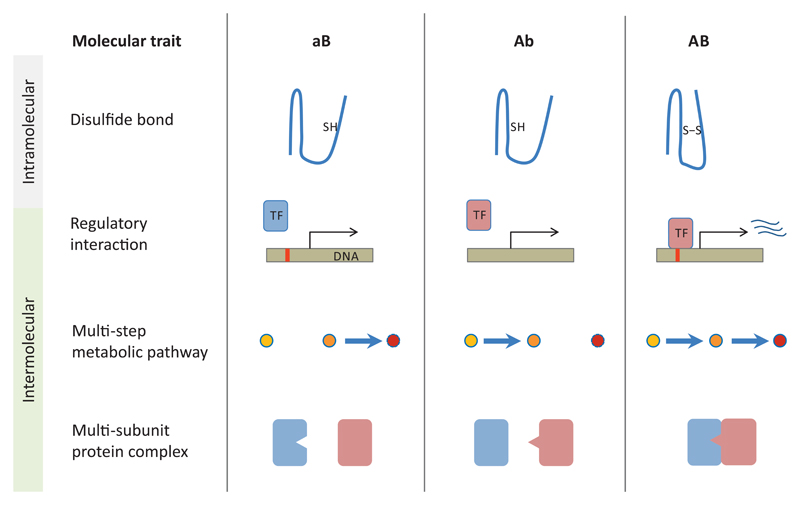
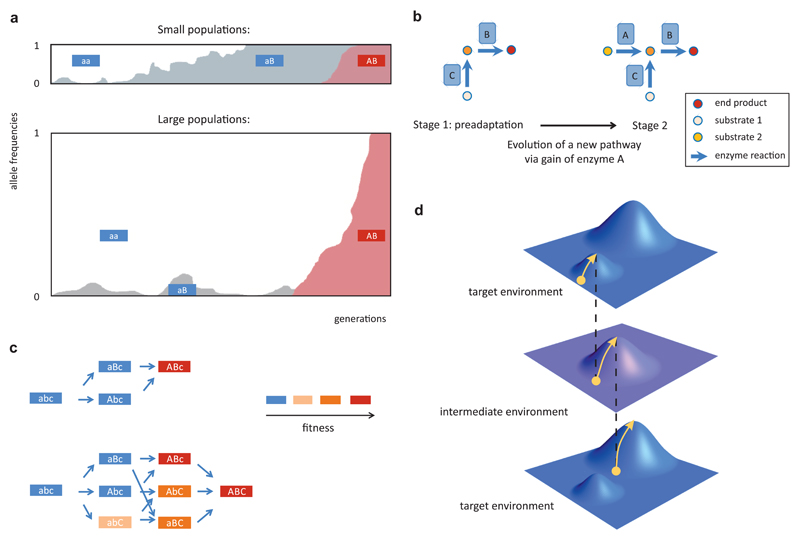
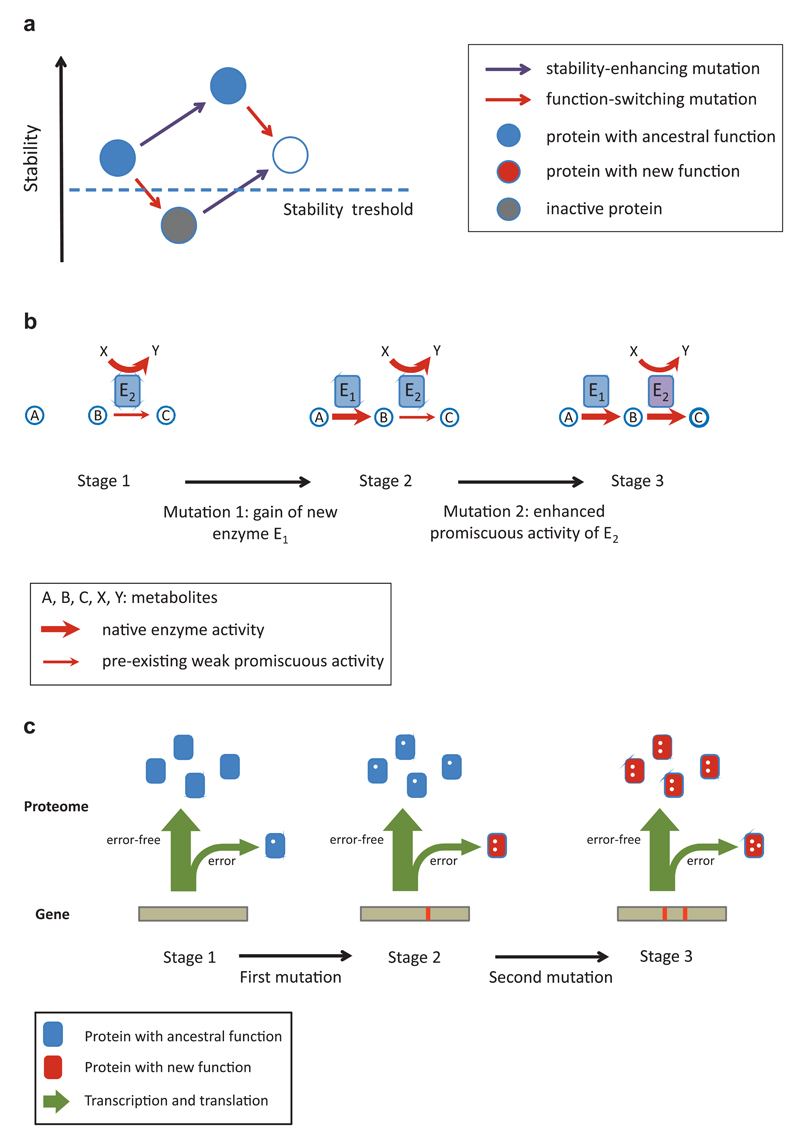
A central challenge in evolutionary biology concerns the mechanisms by which complex adaptations arise. Such adaptations depend on the fixation of multiple, highly specific mutations, where intermediate stages of evolution seemingly provide little or no benefit. It is generally assumed that the establishment of complex adaptations is very slow in nature, as evolution of such traits demands special population genetic or environmental circumstances. However, blueprints of complex adaptations in molecular systems are pervasive, indicating that they can readily evolve. We discuss the prospects and limitations of non-adaptive scenarios, which assume multiple neutral or deleterious steps in the evolution of complex adaptations. Next, we examine how complex adaptations can evolve by natural selection in changing environment. Finally, we argue that molecular 'springboards', such as phenotypic heterogeneity and promiscuous interactions facilitate this process by providing access to new adaptive paths.
Conflict of interest statement
Competing financial interests statement The authors declare no competing financial interests.
Figures
References
-
- Smith JM, Burian R, Kauffman S, Alberch P, et al. Developmental constraints and evolution: a perspective from the Mountain Lake conference on development and evolution. Quarterly Review of Biology. 1985:265–287.
-
- Darwin C. The origin of species. London: Murray; 1859.
-
- Fisher RA. The genetical theory of natural selection: a complete variorum edition. Oxford University Press; 1930.
-
- Wright S. Surfaces of selective value revisited. The American Naturalist. 1988;131:115–123.
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
