

Detection of phasic dopamine by D1 and D2 striatal medium spiny neurons
- PMID: 28782235
- PMCID: PMC5730852
- DOI: 10.1113/JP274475
Detection of phasic dopamine by D1 and D2 striatal medium spiny neurons
Abstract
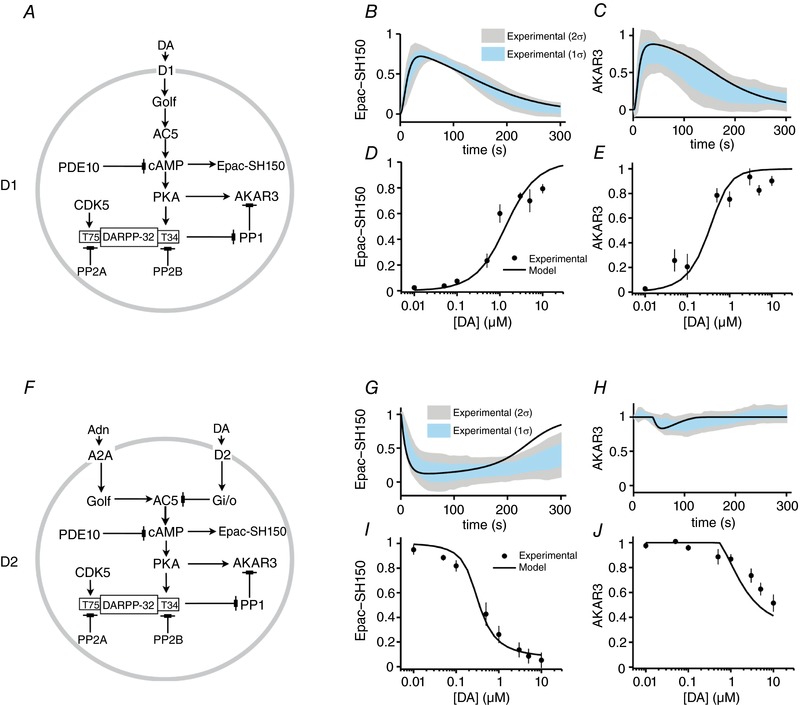
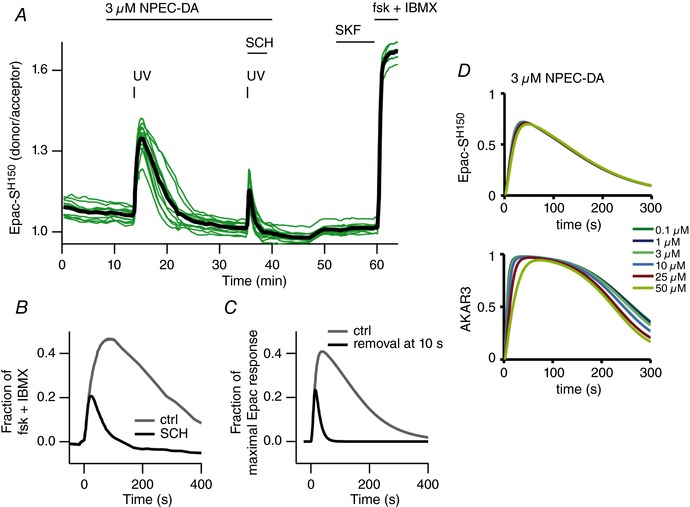
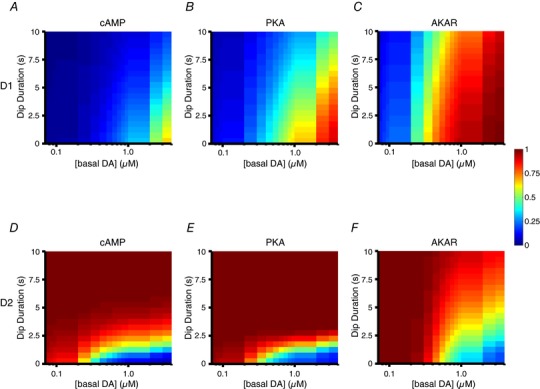
Key points: Brief dopamine events are critical actors of reward-mediated learning in the striatum; the intracellular cAMP-protein kinase A (PKA) response of striatal medium spiny neurons to such events was studied dynamically using a combination of biosensor imaging in mouse brain slices and in silico simulations. Both D1 and D2 medium spiny neurons can sense brief dopamine transients in the sub-micromolar range. While dopamine transients profoundly change cAMP levels in both types of medium spiny neurons, the PKA-dependent phosphorylation level remains unaffected in D2 neurons. At the level of PKA-dependent phosphorylation, D2 unresponsiveness depends on protein phosphatase-1 (PP1) inhibition by DARPP-32. Simulations suggest that D2 medium spiny neurons could detect transient dips in dopamine level.
Abstract: The phasic release of dopamine in the striatum determines various aspects of reward and action selection, but the dynamics of the dopamine effect on intracellular signalling remains poorly understood. We used genetically encoded FRET biosensors in striatal brain slices to quantify the effect of transient dopamine on cAMP or PKA-dependent phosphorylation levels, and computational modelling to further explore the dynamics of this signalling pathway. Medium-sized spiny neurons (MSNs), which express either D1 or D2 dopamine receptors, responded to dopamine by an increase or a decrease in cAMP, respectively. Transient dopamine showed similar sub-micromolar efficacies on cAMP in both D1 and D2 MSNs, thus challenging the commonly accepted notion that dopamine efficacy is much higher on D2 than on D1 receptors. However, in D2 MSNs, the large decrease in cAMP level triggered by transient dopamine did not translate to a decrease in PKA-dependent phosphorylation level, owing to the efficient inhibition of protein phosphatase 1 by DARPP-32. Simulations further suggested that D2 MSNs can also operate in a 'tone-sensing' mode, allowing them to detect transient dips in basal dopamine. Overall, our results show that D2 MSNs may sense much more complex patterns of dopamine than previously thought.
Keywords: biosensor imaging; dopamine; intracellular second messengers.
© 2017 The Authors. The Journal of Physiology © 2017 The Physiological Society.
Figures
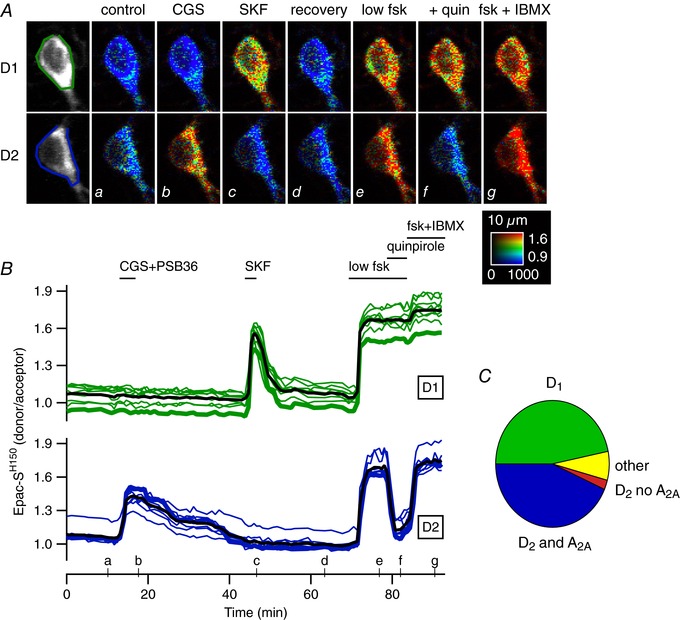
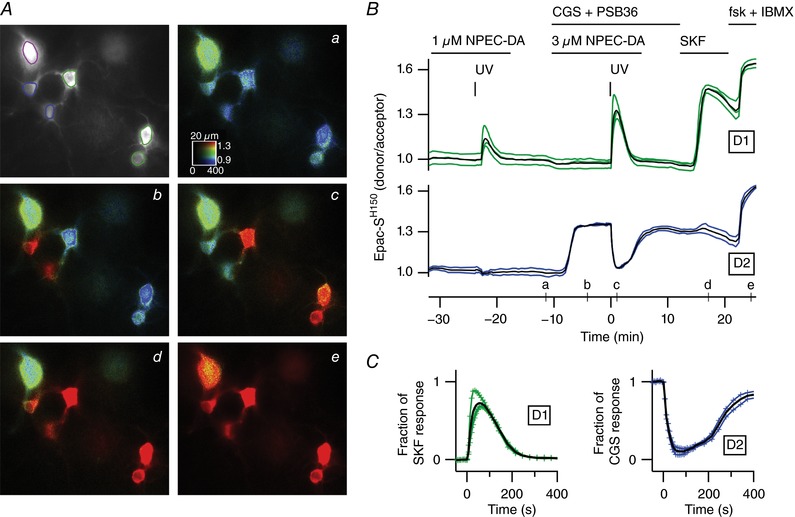
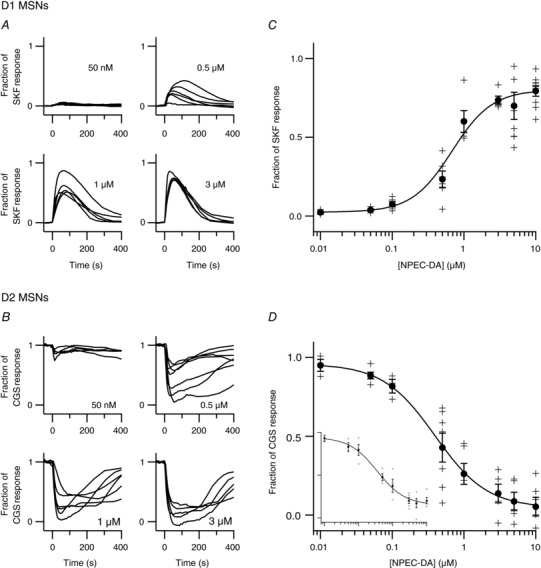
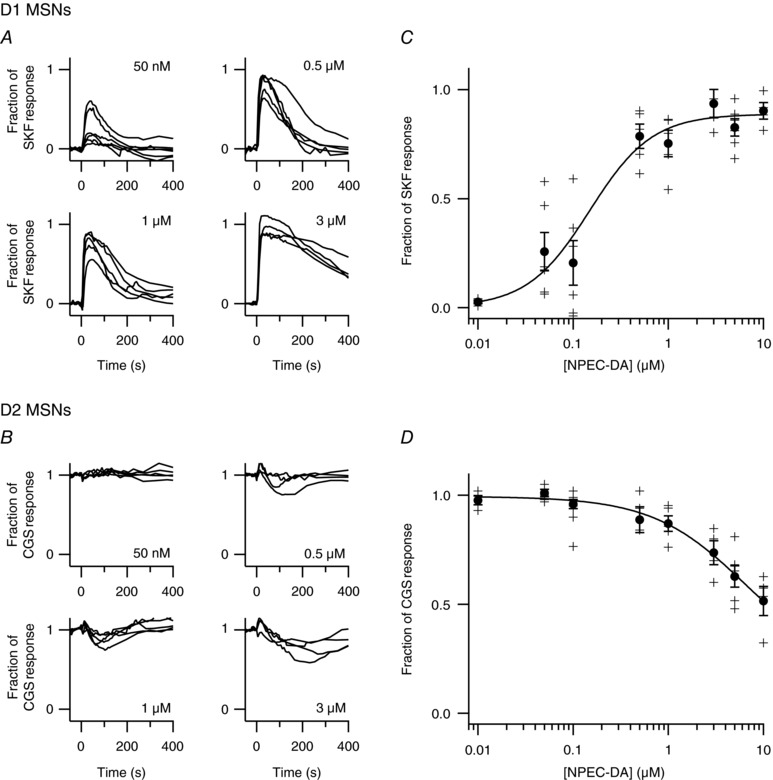

Comment in
-
Striatal neurons get a kick out of dopamine.J Physiol. 2017 Dec 15;595(24):7271-7272. doi: 10.1113/JP275079. Epub 2017 Nov 19. J Physiol. 2017. PMID: 29105113 Free PMC article. No abstract available.
References
-
- Albert PR, Neve KA, Bunzow JR & Civelli O (1990). Coupling of a cloned rat dopamine‐D2 receptor to inhibition of adenylyl cyclase and prolactin secretion. J Biol Chem 265, 2098–2104. - PubMed
-
- Allen MD & Zhang J (2006). Subcellular dynamics of protein kinase A activity visualized by FRET‐based reporters. Biochem Biophys Res Commun 348, 716–721. - PubMed
-
- Arbuthnott GW & Wickens J (2007). Space, time and dopamine. Trends Neurosci 30, 62–69. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases