

Katanin spiral and ring structures shed light on power stroke for microtubule severing
- PMID: 28783150
- PMCID: PMC7152510
- DOI: 10.1038/nsmb.3448
Katanin spiral and ring structures shed light on power stroke for microtubule severing
Abstract
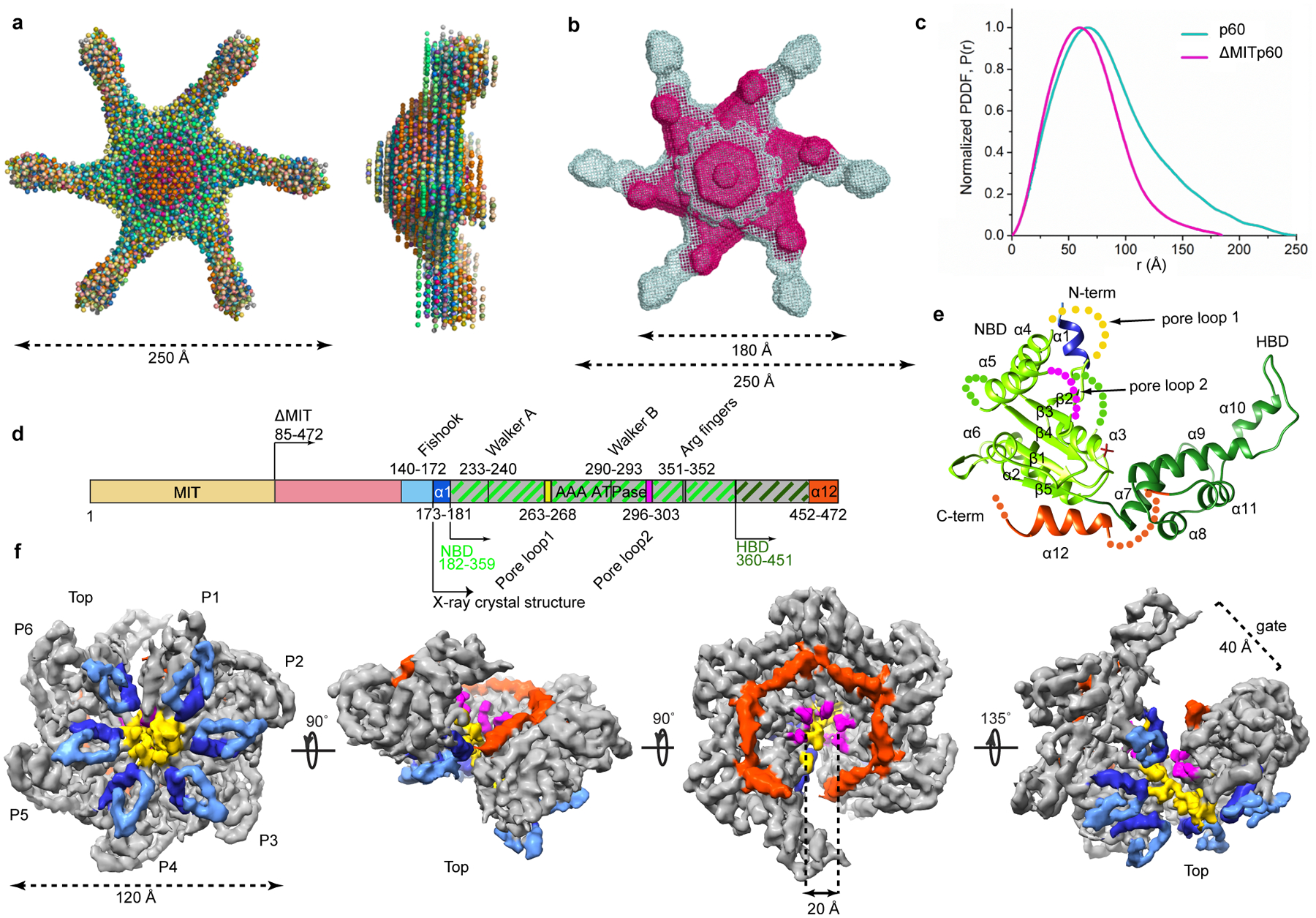
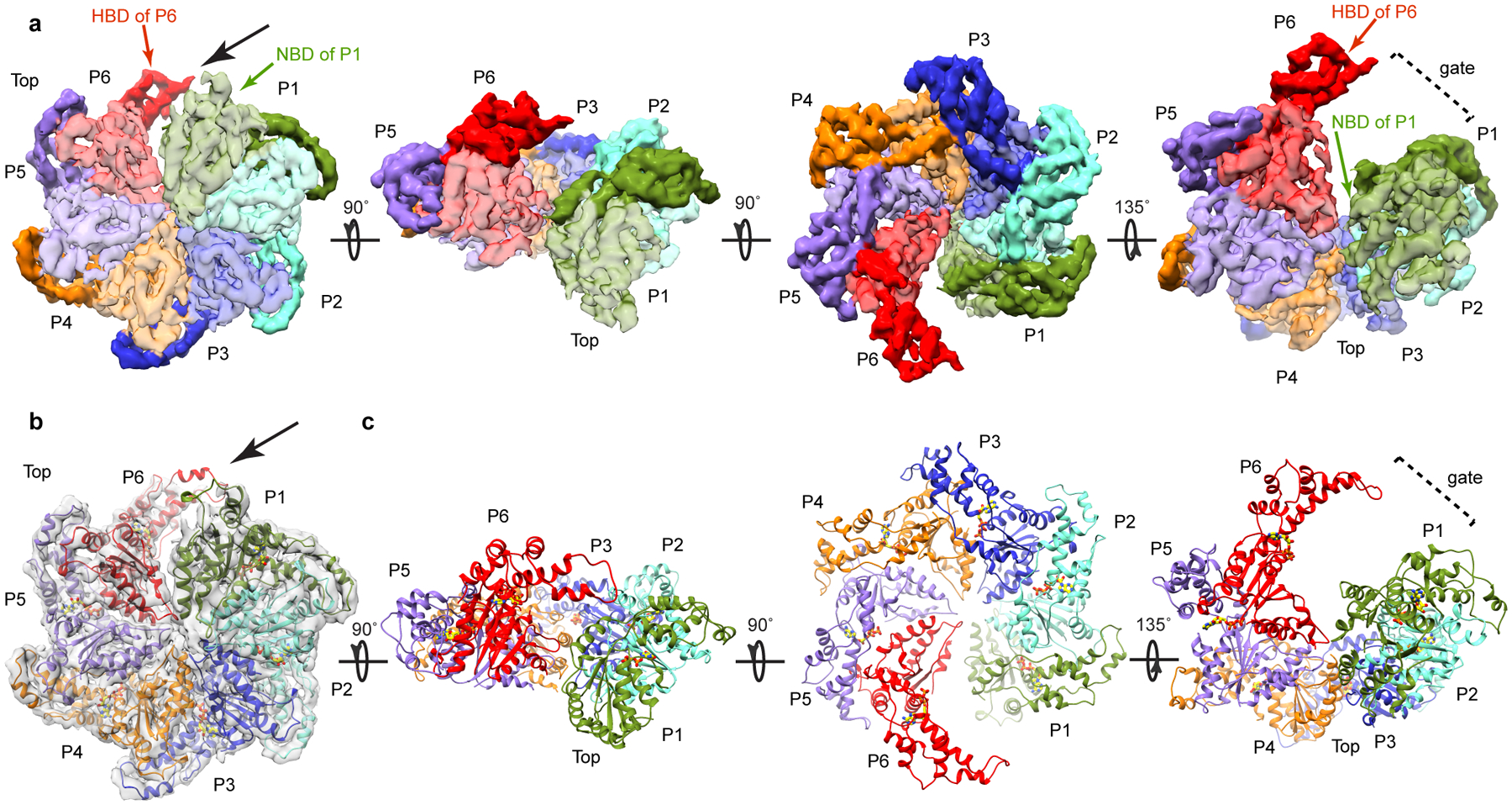
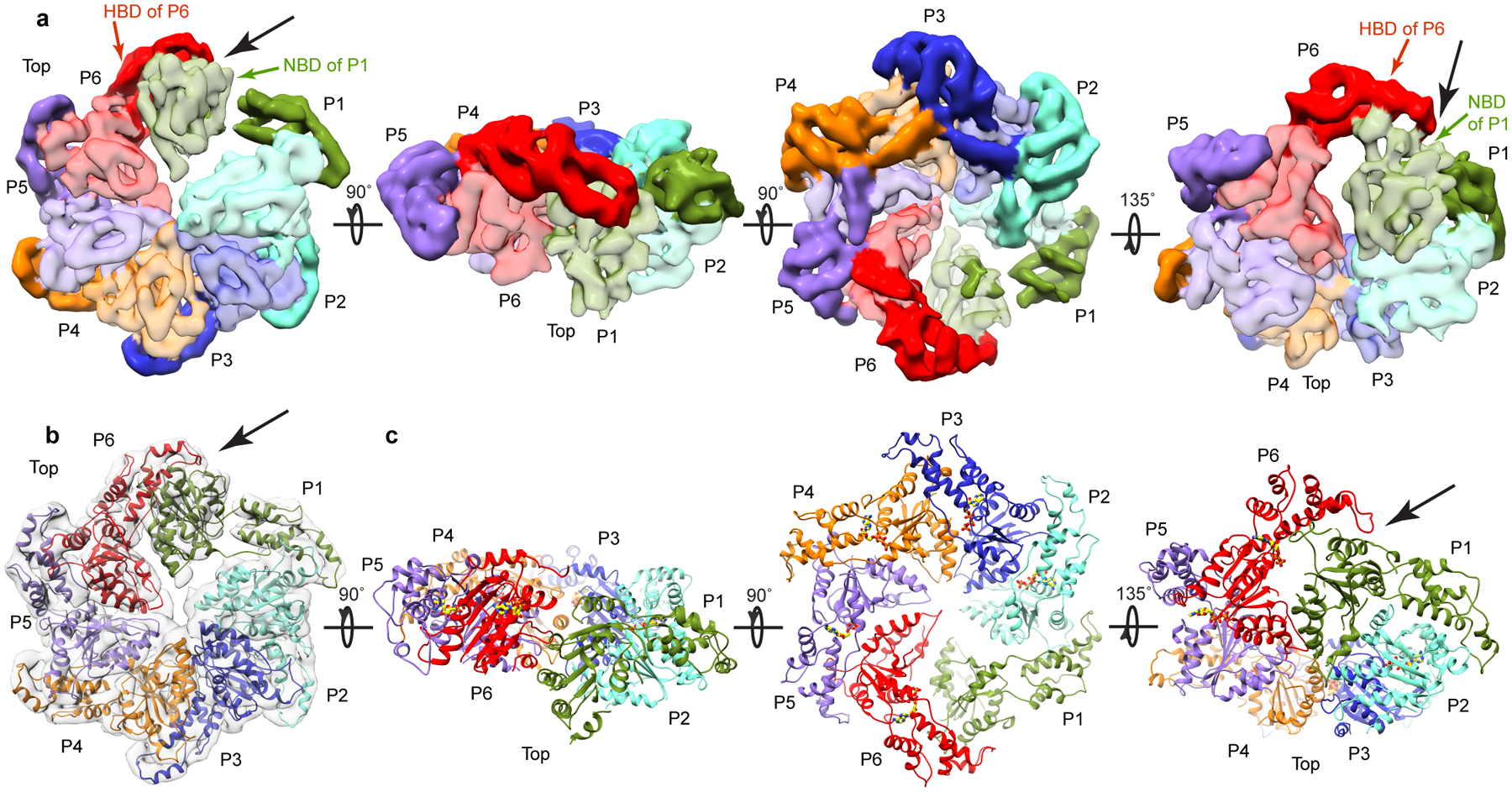
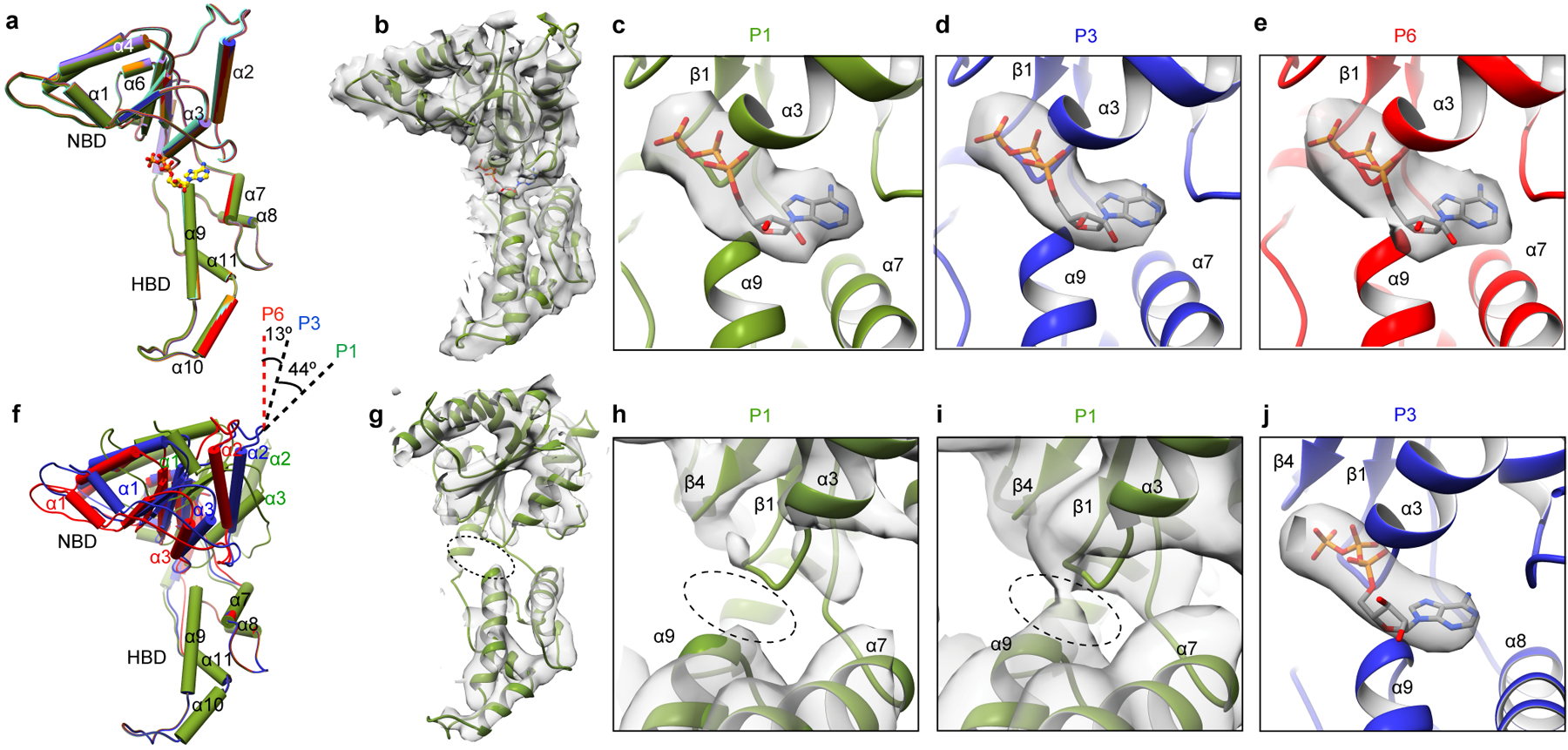
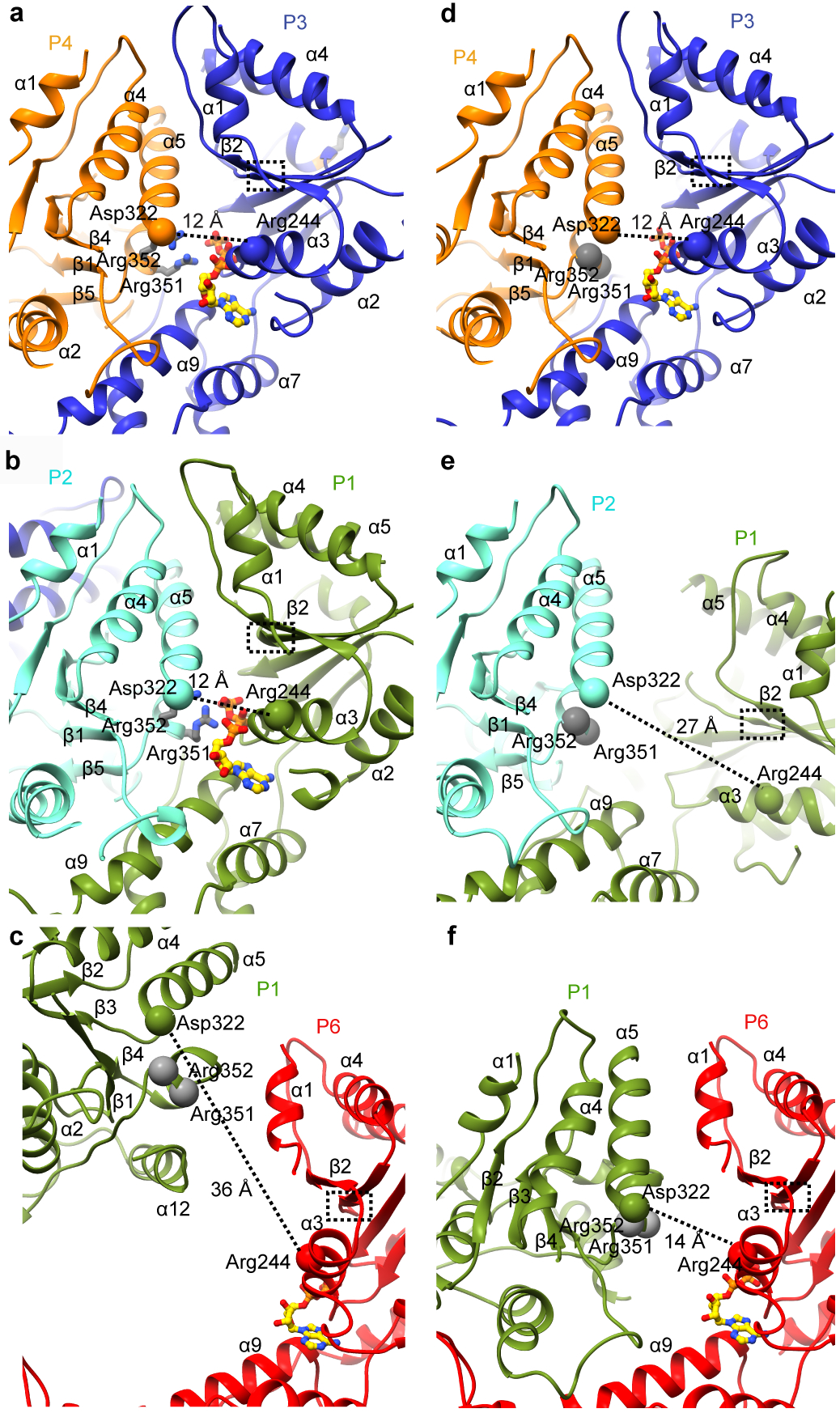
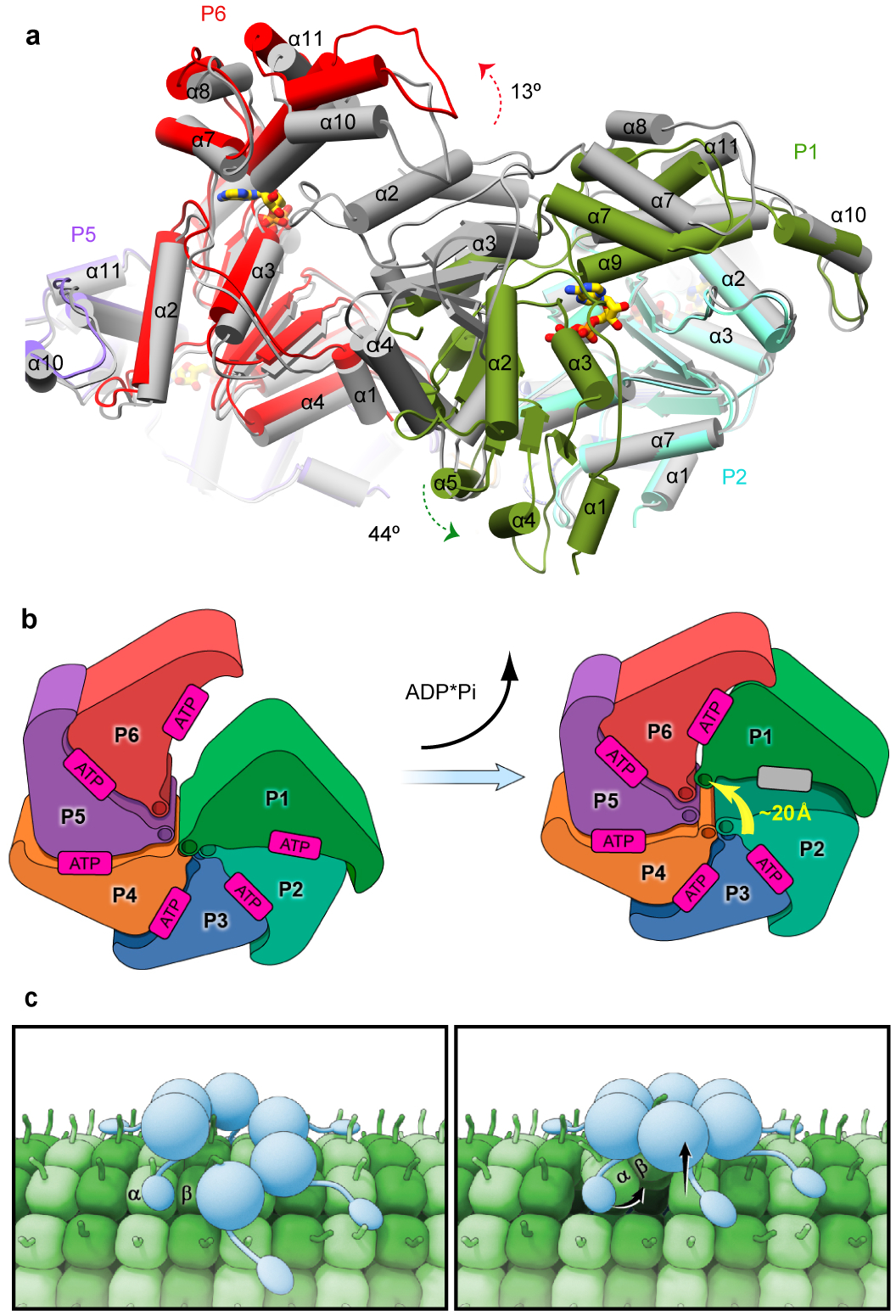
Microtubule-severing enzymes katanin, spastin and fidgetin are AAA ATPases important for the biogenesis and maintenance of complex microtubule arrays in axons, spindles and cilia. Because of a lack of known 3D structures for these enzymes, their mechanism of action has remained poorly understood. Here we report the X-ray crystal structure of the monomeric AAA katanin module from Caenorhabditis elegans and cryo-EM reconstructions of the hexamer in two conformations. The structures reveal an unexpected asymmetric arrangement of the AAA domains mediated by structural elements unique to microtubule-severing enzymes and critical for their function. The reconstructions show that katanin cycles between open spiral and closed ring conformations, depending on the ATP occupancy of a gating protomer that tenses or relaxes interprotomer interfaces. Cycling of the hexamer between these conformations would provide the power stroke for microtubule severing.
Conflict of interest statement
COMPETING FINANCIAL INTERESTS
The authors declare no competing financial interests.
Figures
Similar articles
-
Structural basis for disassembly of katanin heterododecamers.J Biol Chem. 2018 Jul 6;293(27):10590-10605. doi: 10.1074/jbc.RA117.001215. Epub 2018 May 11. J Biol Chem. 2018. PMID: 29752405 Free PMC article.
-
Katanin Grips the β-Tubulin Tail through an Electropositive Double Spiral to Sever Microtubules.Dev Cell. 2020 Jan 6;52(1):118-131.e6. doi: 10.1016/j.devcel.2019.10.010. Epub 2019 Nov 14. Dev Cell. 2020. PMID: 31735665 Free PMC article.
-
The spindle assembly function of Caenorhabditis elegans katanin does not require microtubule-severing activity.Mol Biol Cell. 2011 May;22(9):1550-60. doi: 10.1091/mbc.E10-12-0951. Epub 2011 Mar 3. Mol Biol Cell. 2011. PMID: 21372175 Free PMC article.
-
Meiotic Clade AAA ATPases: Protein Polymer Disassembly Machines.J Mol Biol. 2016 May 8;428(9 Pt B):1897-911. doi: 10.1016/j.jmb.2015.11.004. Epub 2015 Nov 10. J Mol Biol. 2016. PMID: 26555750 Free PMC article. Review.
-
[Microtubule severing proteins - structure and activity regulation].Postepy Biochem. 2016;62(1):46-51. Postepy Biochem. 2016. PMID: 28132444 Review. Polish.
Cited by
-
CsKTN1 for a katanin p60 subunit is associated with the regulation of fruit elongation in cucumber (Cucumis sativus L.).Theor Appl Genet. 2021 Aug;134(8):2429-2441. doi: 10.1007/s00122-021-03833-y. Epub 2021 May 27. Theor Appl Genet. 2021. PMID: 34043036
-
Cryo-EM structure of the essential ribosome assembly AAA-ATPase Rix7.Nat Commun. 2019 Jan 31;10(1):513. doi: 10.1038/s41467-019-08373-0. Nat Commun. 2019. PMID: 30705282 Free PMC article.
-
Structure and mechanism of the ESCRT pathway AAA+ ATPase Vps4.Biochem Soc Trans. 2019 Feb 28;47(1):37-45. doi: 10.1042/BST20180260. Epub 2019 Jan 15. Biochem Soc Trans. 2019. PMID: 30647138 Free PMC article. Review.
-
The katanin A-subunits KATNA1 and KATNAL1 act co-operatively in mammalian meiosis and spermiogenesis to achieve male fertility.Development. 2023 Nov 15;150(22):dev201956. doi: 10.1242/dev.201956. Epub 2023 Nov 13. Development. 2023. PMID: 37882691 Free PMC article.
-
Spastin accumulation and motor neuron defects caused by a novel SPAST splice site mutation.J Transl Med. 2024 Sep 27;22(1):872. doi: 10.1186/s12967-024-05669-8. J Transl Med. 2024. PMID: 39334479 Free PMC article.
References
-
- Zhang Q, Fishel E, Bertroche T & Dixit R Microtubule severing at crossover sites by katanin generates ordered cortical microtubule arrays in Arabidopsis. Current Biology 23, 2191–2195 (2013). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
