

Glioblastoma stem cells exploit the αvβ8 integrin-TGFβ1 signaling axis to drive tumor initiation and progression
- PMID: 28783169
- PMCID: PMC5882487
- DOI: 10.1038/onc.2017.248
Glioblastoma stem cells exploit the αvβ8 integrin-TGFβ1 signaling axis to drive tumor initiation and progression
Abstract
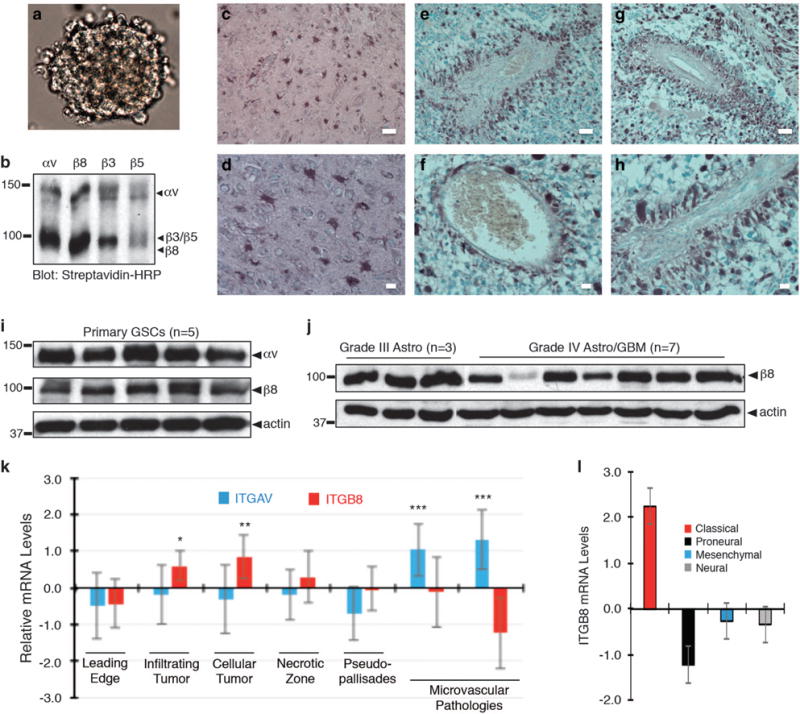
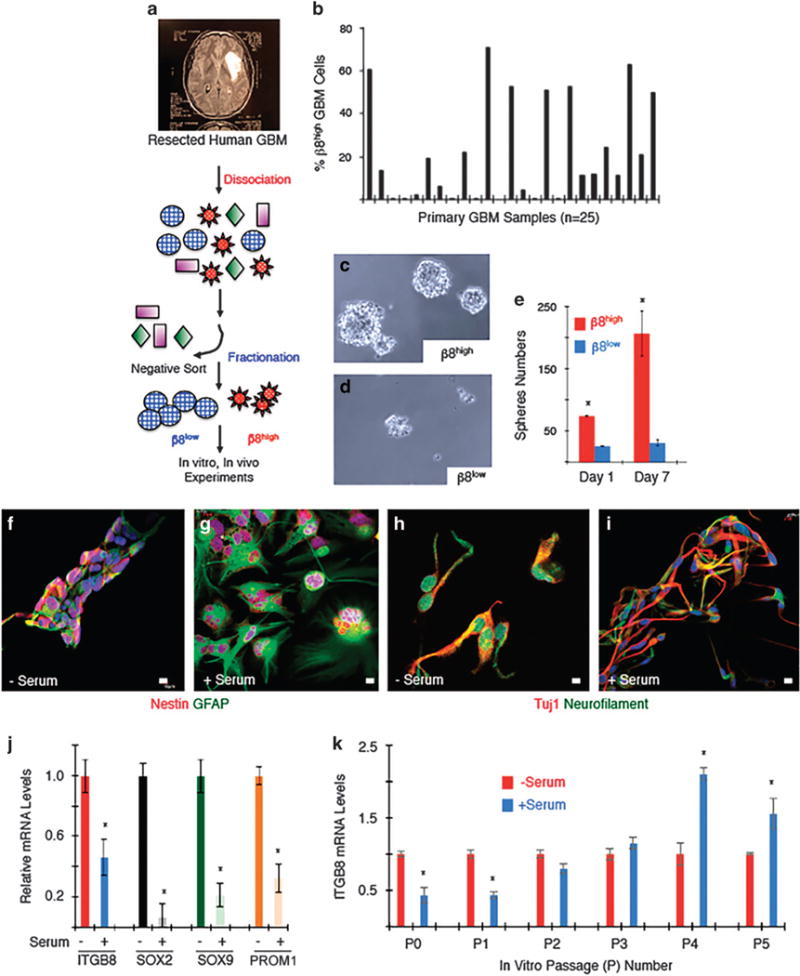
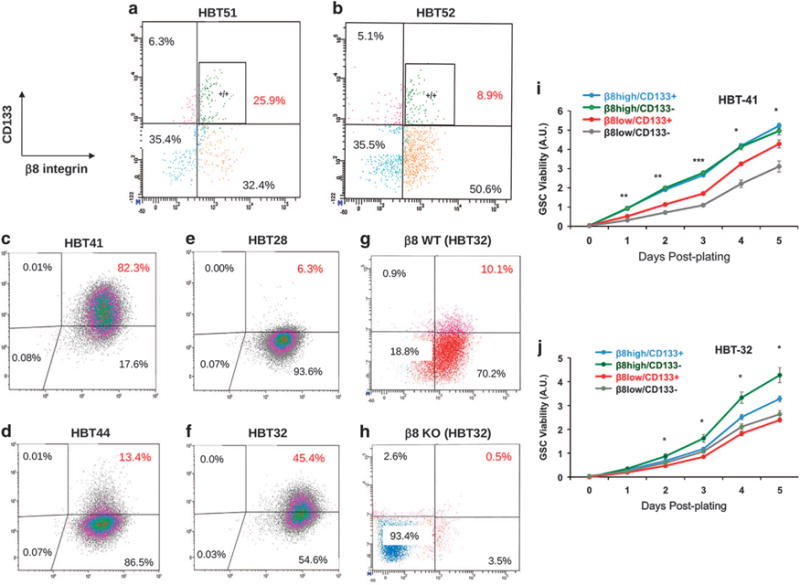
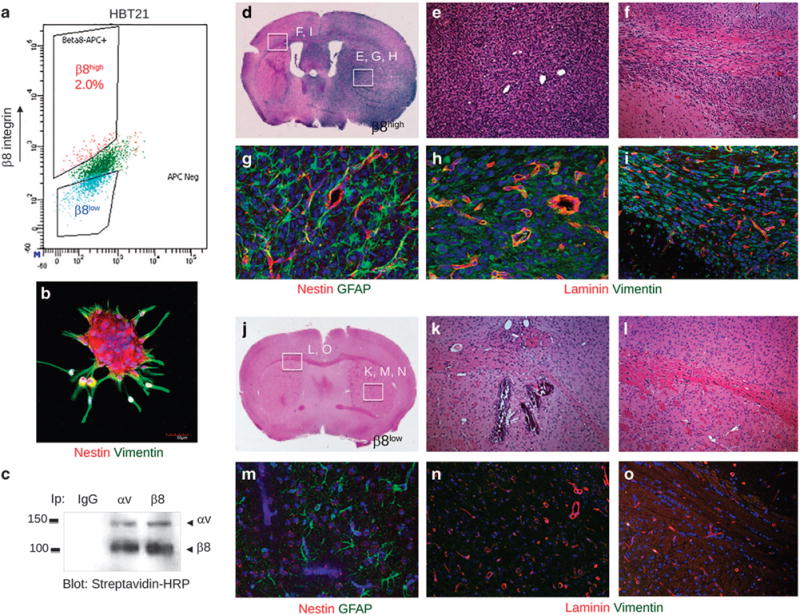
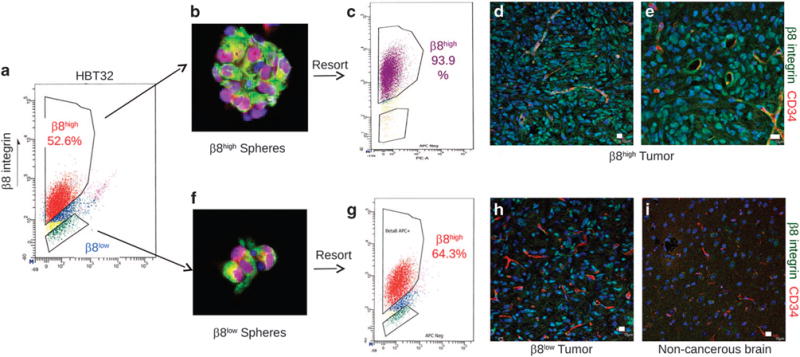
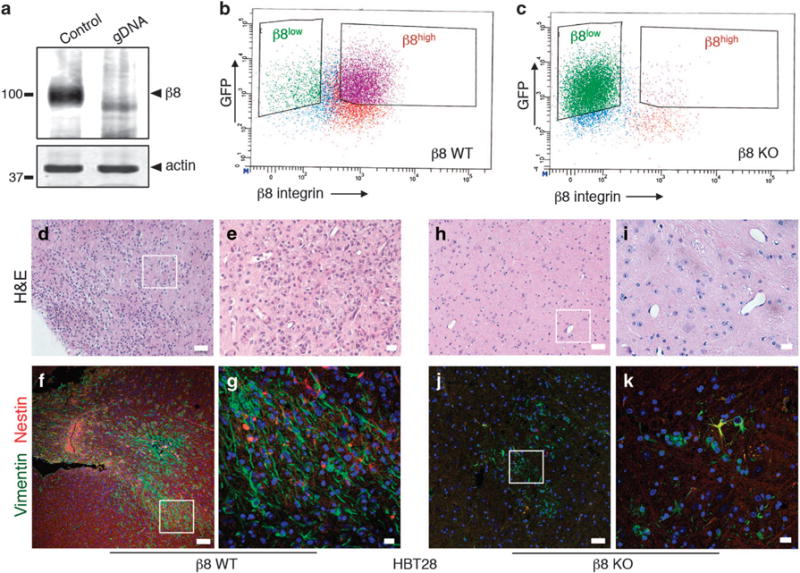
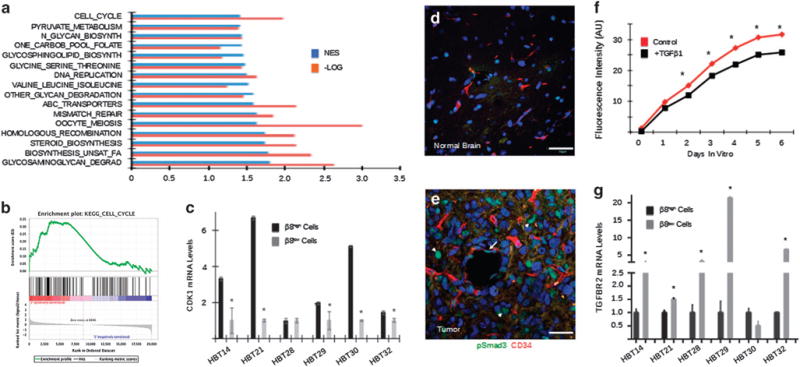
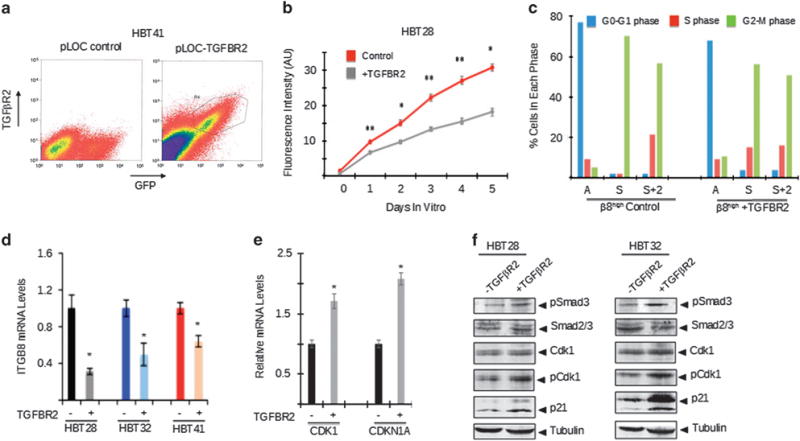
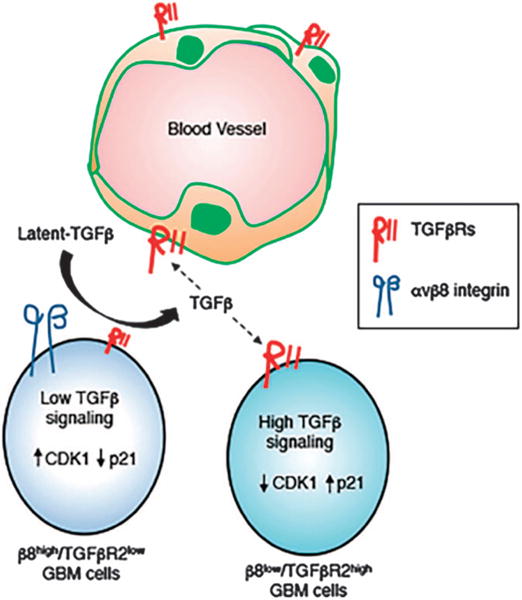
Glioblastoma (GBM) is a primary brain cancer that contains populations of stem-like cancer cells (GSCs) that home to specialized perivascular niches. GSC interactions with their niche influence self-renewal, differentiation and drug resistance, although the pathways underlying these events remain largely unknown. Here, we report that the integrin αvβ8 and its latent transforming growth factor β1 (TGFβ1) protein ligand have central roles in promoting niche co-option and GBM initiation. αvβ8 integrin is highly expressed in GSCs and is essential for self-renewal and lineage commitment in vitro. Fractionation of β8high cells from freshly resected human GBM samples also reveals a requirement for this integrin in tumorigenesis in vivo. Whole-transcriptome sequencing reveals that αvβ8 integrin regulates tumor development, in part, by driving TGFβ1-induced DNA replication and mitotic checkpoint progression. Collectively, these data identify the αvβ8 integrin-TGFβ1 signaling axis as crucial for exploitation of the perivascular niche and identify potential therapeutic targets for inhibiting tumor growth and progression in patients with GBM.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Engagement of cellular prion protein with the co-chaperone Hsp70/90 organizing protein regulates the proliferation of glioblastoma stem-like cells.Stem Cell Res Ther. 2017 Apr 17;8(1):76. doi: 10.1186/s13287-017-0518-1. Stem Cell Res Ther. 2017. PMID: 28412969 Free PMC article.
-
αvβ8 integrin interacts with RhoGDI1 to regulate Rac1 and Cdc42 activation and drive glioblastoma cell invasion.Mol Biol Cell. 2013 Feb;24(4):474-82. doi: 10.1091/mbc.E12-07-0521. Epub 2013 Jan 2. Mol Biol Cell. 2013. PMID: 23283986 Free PMC article.
-
A Novel Role of BIRC3 in Stemness Reprogramming of Glioblastoma.Int J Mol Sci. 2021 Dec 28;23(1):297. doi: 10.3390/ijms23010297. Int J Mol Sci. 2021. PMID: 35008722 Free PMC article.
-
Glioblastoma Stem Cells at the Nexus of Tumor Heterogeneity, Immune Evasion, and Therapeutic Resistance.Cells. 2025 Apr 9;14(8):562. doi: 10.3390/cells14080562. Cells. 2025. PMID: 40277888 Free PMC article. Review.
-
Biomimetic models to examine microenvironmental regulation of glioblastoma stem cells.Cancer Lett. 2018 Aug 10;429:41-53. doi: 10.1016/j.canlet.2018.05.007. Epub 2018 May 8. Cancer Lett. 2018. PMID: 29746930 Review.
Cited by
-
Integrins regulate stemness in solid tumor: an emerging therapeutic target.J Hematol Oncol. 2021 Oct 29;14(1):177. doi: 10.1186/s13045-021-01192-1. J Hematol Oncol. 2021. PMID: 34715893 Free PMC article. Review.
-
The Role of Hypoxia and Cancer Stem Cells in Development of Glioblastoma.Cancers (Basel). 2023 May 4;15(9):2613. doi: 10.3390/cancers15092613. Cancers (Basel). 2023. PMID: 37174078 Free PMC article. Review.
-
Mechanical Constraints in Tumor Guide Emergent Spatial Patterns of Glioblastoma Cancer Stem Cells.Mechanobiol Med. 2024 Mar;2(1):100027. doi: 10.1016/j.mbm.2023.100027. Epub 2023 Oct 29. Mechanobiol Med. 2024. PMID: 38770108 Free PMC article.
-
Integrins in Cancer Drug Resistance: Molecular Mechanisms and Clinical Implications.Int J Mol Sci. 2025 Mar 28;26(7):3143. doi: 10.3390/ijms26073143. Int J Mol Sci. 2025. PMID: 40243917 Free PMC article. Review.
-
IFITM3 promotes glioblastoma stem cell-mediated angiogenesis via regulating JAK/STAT3/bFGF signaling pathway.Cell Death Dis. 2024 Jan 13;15(1):45. doi: 10.1038/s41419-023-06416-5. Cell Death Dis. 2024. PMID: 38218875 Free PMC article.
References
-
- Sanai N, Alvarez-Buylla A, Berger MS. Neural stem cells and the origin of gliomas. N Engl J Med. 2005;353:811–822. - PubMed
-
- Bao S, Wu Q, McLendon RE, Hao Y, Shi Q, Hjelmeland AB, et al. Glioma stem cells promote radioresistance by preferential activation of the DNA damage response. Nature. 2006;444:756–760. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
