

A Noncanonical Binding Site in the EVH1 Domain of Vasodilator-Stimulated Phosphoprotein Regulates Its Interactions with the Proline Rich Region of Zyxin
- PMID: 28783324
- PMCID: PMC5724578
- DOI: 10.1021/acs.biochem.7b00618
A Noncanonical Binding Site in the EVH1 Domain of Vasodilator-Stimulated Phosphoprotein Regulates Its Interactions with the Proline Rich Region of Zyxin
Abstract
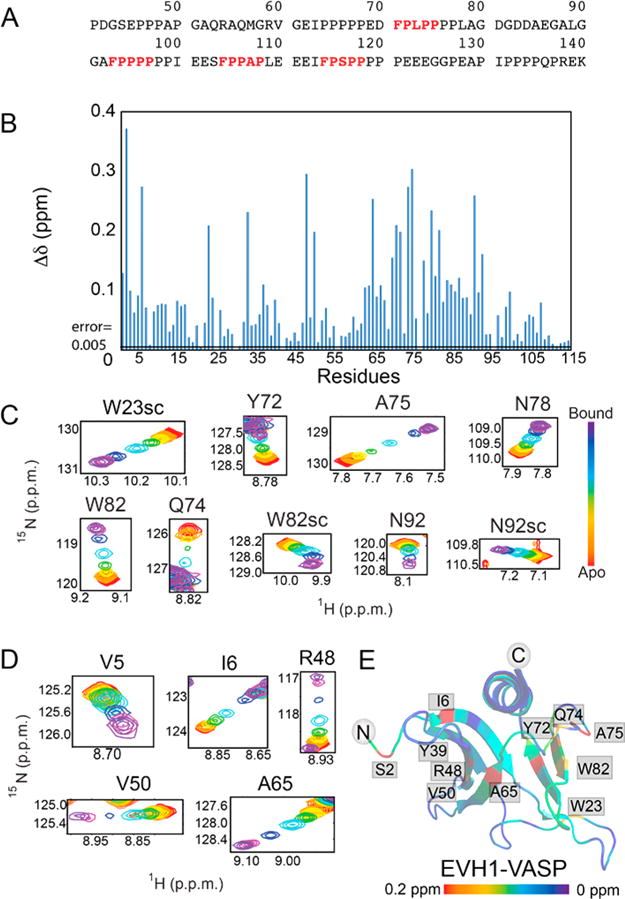
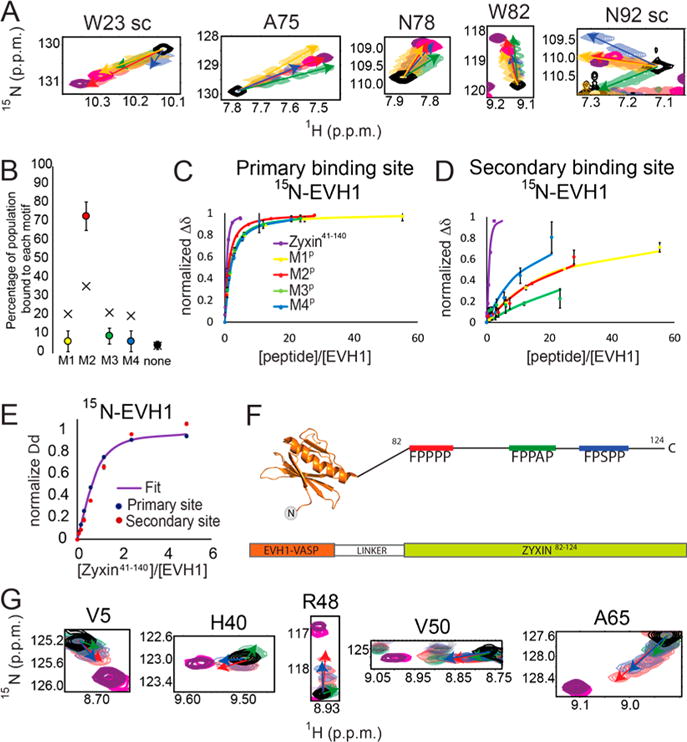
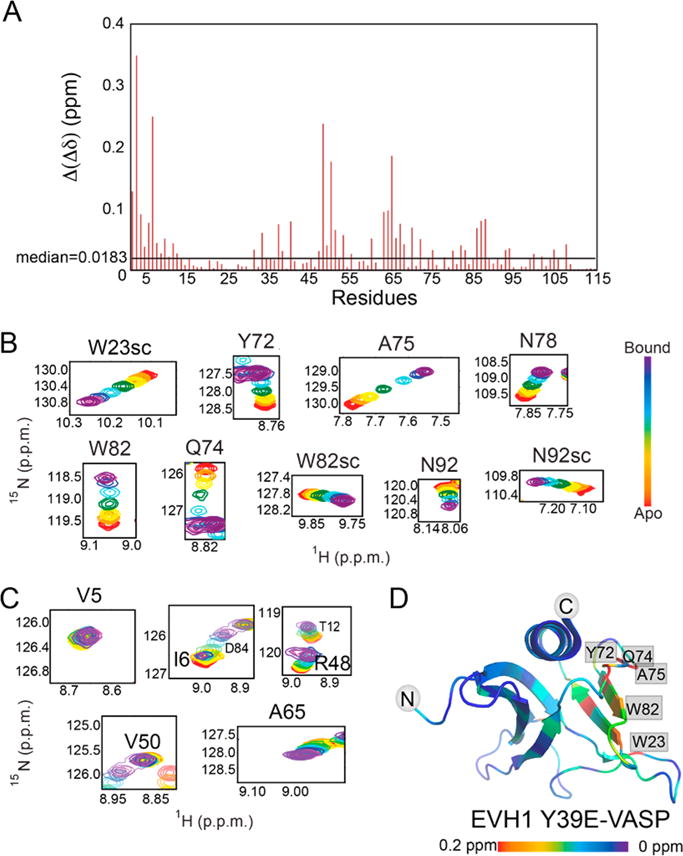
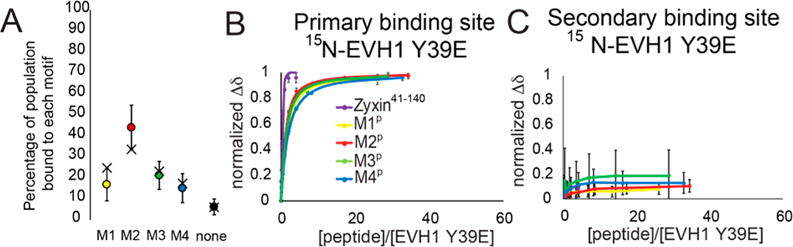
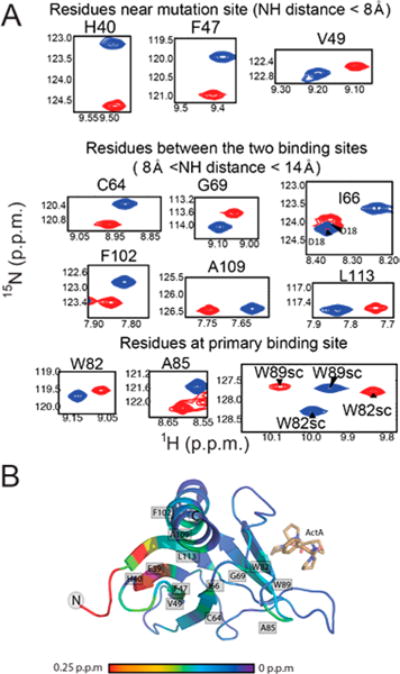
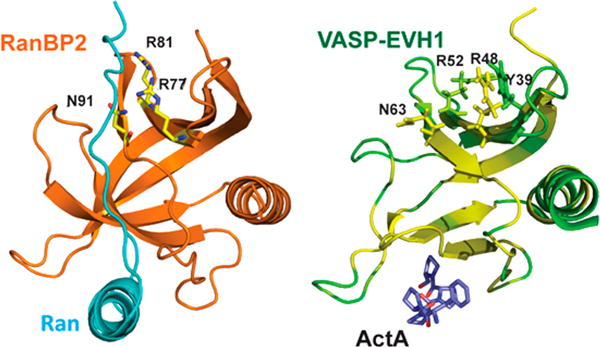
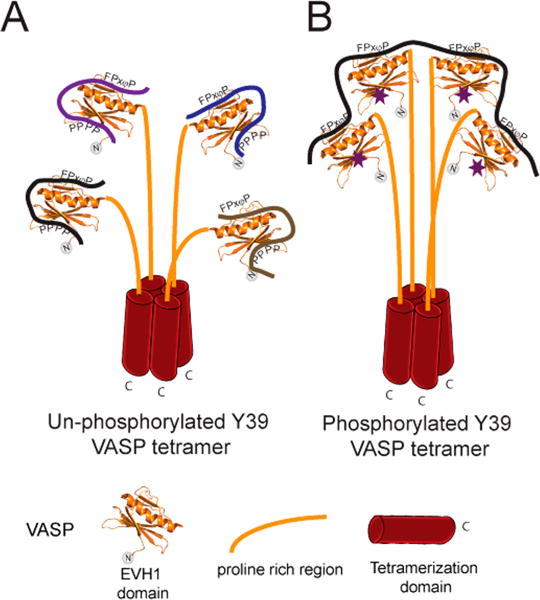
Vasodilator-stimulated phosphoprotein (VASP) is a processive actin polymerase with roles in the control of cell shape and cell migration. Through interaction with the cytoskeletal adaptor protein Zyxin, VASP can localize to damaged stress fibers where it serves to repair and reinforce these structures. VASP localization is mediated by its N-terminal Ena/VASP homology (EVH1) domain, which binds to the (W/F)PxφP motif (most commonly occurring as FPPPP) found in cytoskeletal proteins such as vinculin, lamellipodin, and Zyxin. Sequentially close clusters of four or five of these motifs frequently occur, as in the proline rich region of Zyxin with four such motifs. This suggests that tetrameric VASP might bind very tightly to Zyxin through avidity, with all four EVH1 domains binding to a single Zyxin molecule. Here, quantitative nuclear magnetic resonance titration analysis reveals a dominant bivalent 1:1 (Zyxin:EVH1) interaction between the Zyxin proline rich region and the VASP EVH1 domain that utilizes the EVH1 canonical binding site and a novel secondary binding site on the opposite face of the EVH1 domain. We further show that binding to the secondary binding site is specifically inhibited by mutation of VASP EVH1 domain residue Y39 to E, which mimics Abl-induced phosphorylation of Y39. On the basis of these findings, we propose a model in which phosphorylation of Y39 acts as a stoichiometry switch that governs binding partner selection by the constitutive VASP tetramer. These results have broader implications for other multivalent VASP EVH1 domain binding partners and for furthering our understanding of the role of Y39 phosphorylation in regulating VASP localization and cellular function.
Conflict of interest statement
Theauthors declare no competing financial interest.
Figures
References
-
- Krause M, Dent EW, Bear JE, Loureiro JJ, Gertler FB. Ena/VASP proteins: regulators of the actin cytoskeleton and cell migration. Annu Rev Cell Dev Biol. 2003;19:541–564. - PubMed
-
- Bear JE, Svitkina TM, Krause M, Schafer DA, Loureiro JJ, Strasser GA, Maly IV, Chaga OY, Cooper JA, Borisy GG, Gertler FB. Antagonism between Ena/VASP proteins and actin filament capping regulates fibroblast motility. Cell. 2002;109:509–521. - PubMed
-
- Lebrand C, Dent EW, Strasser GA, Lanier LM, Krause M, Svitkina TM, Borisy GG, Gertler FB. Critical role of Ena/VASP proteins for filopodia formation in neurons and in function downstream of netrin-1. Neuron. 2004;42:37–49. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
