

The Three-Dimensional Organization of Mammalian Genomes
- PMID: 28783961
- PMCID: PMC5837811
- DOI: 10.1146/annurev-cellbio-100616-060531
The Three-Dimensional Organization of Mammalian Genomes
Abstract
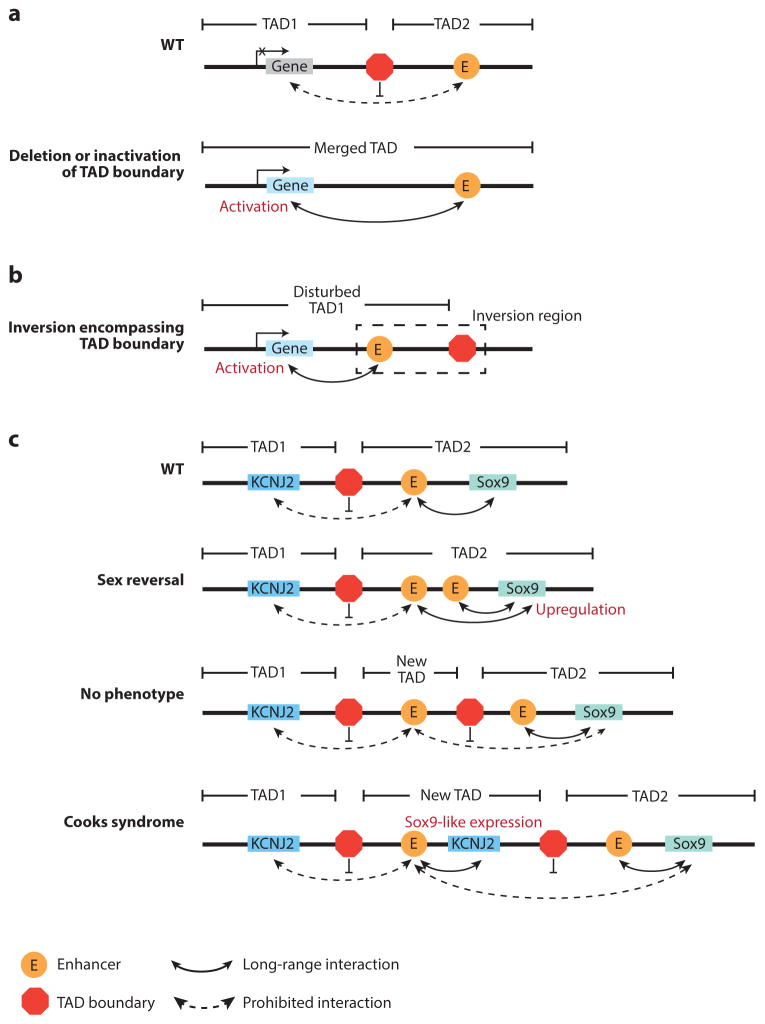
Animal development depends on not only the linear genome sequence that embeds millions of cis-regulatory elements, but also the three-dimensional (3D) chromatin architecture that orchestrates the interplay between cis-regulatory elements and their target genes. Compared to our knowledge of the cis-regulatory sequences, the understanding of the 3D genome organization in human and other eukaryotes is still limited. Recent advances in technologies to map the 3D genome architecture have greatly accelerated the pace of discovery. Here, we review emerging concepts of chromatin organization in mammalian cells, discuss the dynamics of chromatin conformation during development, and highlight important roles for chromatin organization in cancer and other human diseases.
Keywords: 3D chromatin organization; TAD; chromosome conformation capture; gene regulation; topologically associating domain.
Figures
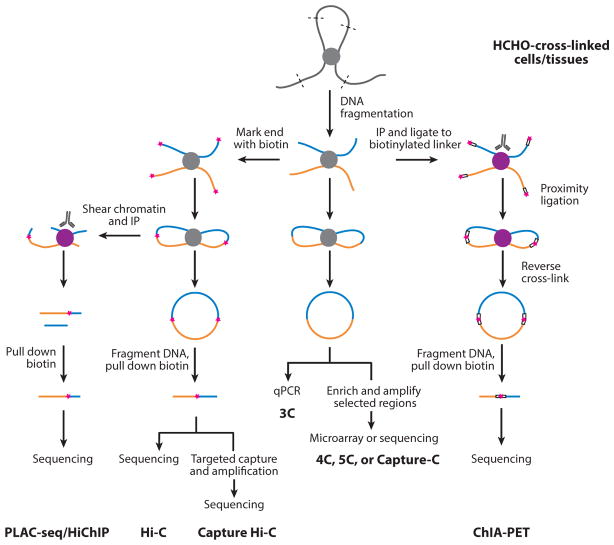
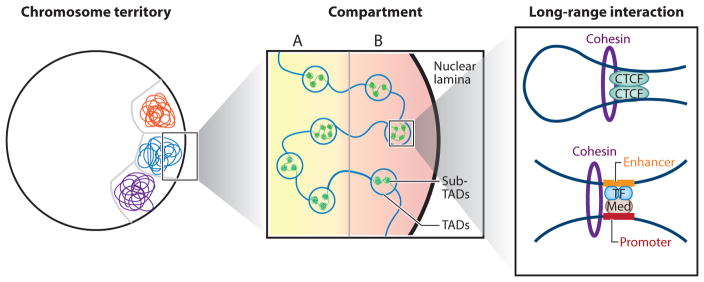
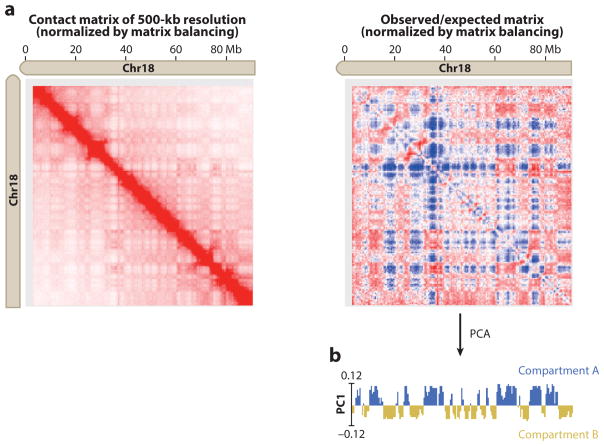
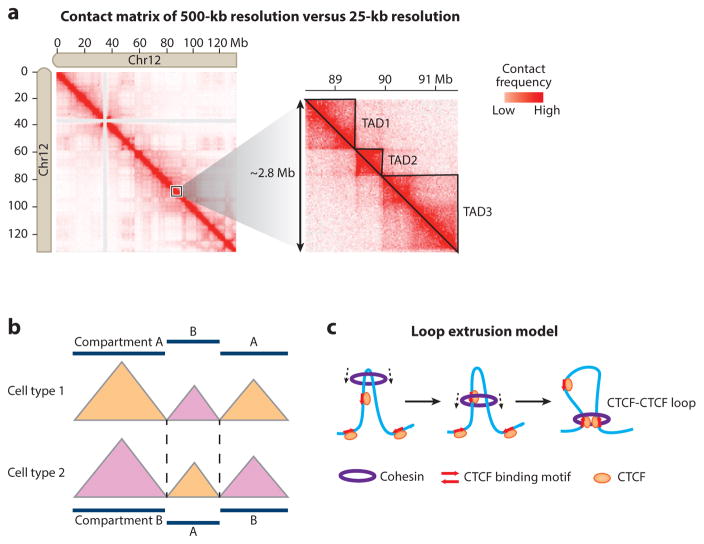
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
