

Identification of the Tau phosphorylation pattern that drives its aggregation
- PMID: 28784767
- PMCID: PMC5576827
- DOI: 10.1073/pnas.1708448114
Identification of the Tau phosphorylation pattern that drives its aggregation
Abstract
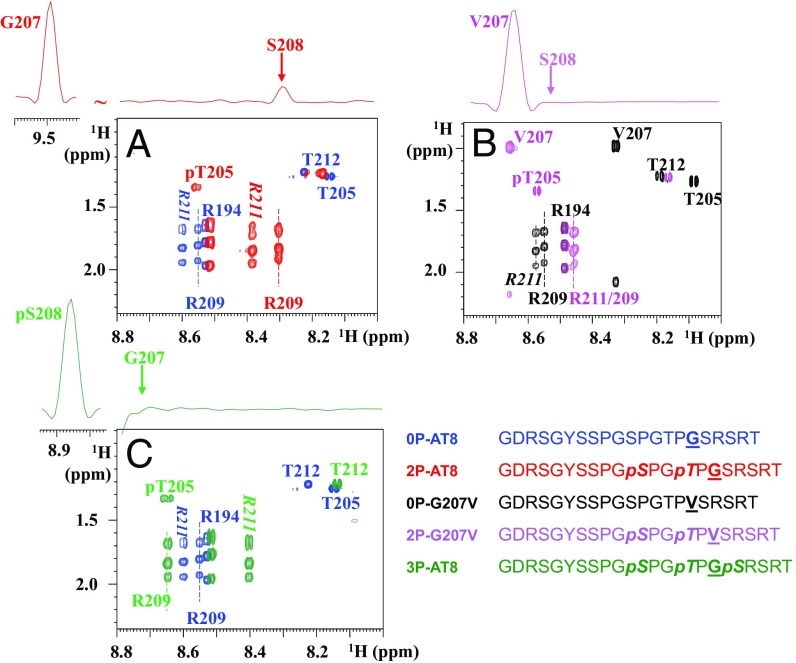
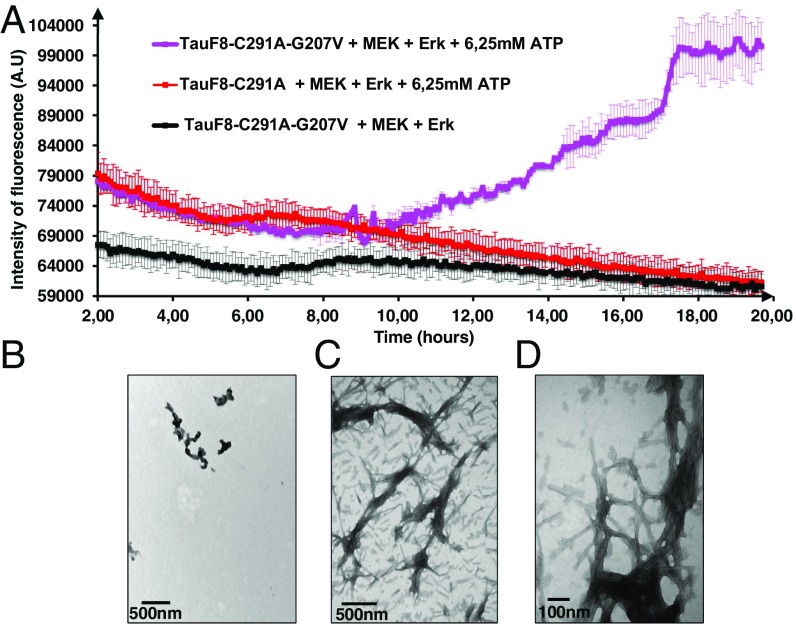
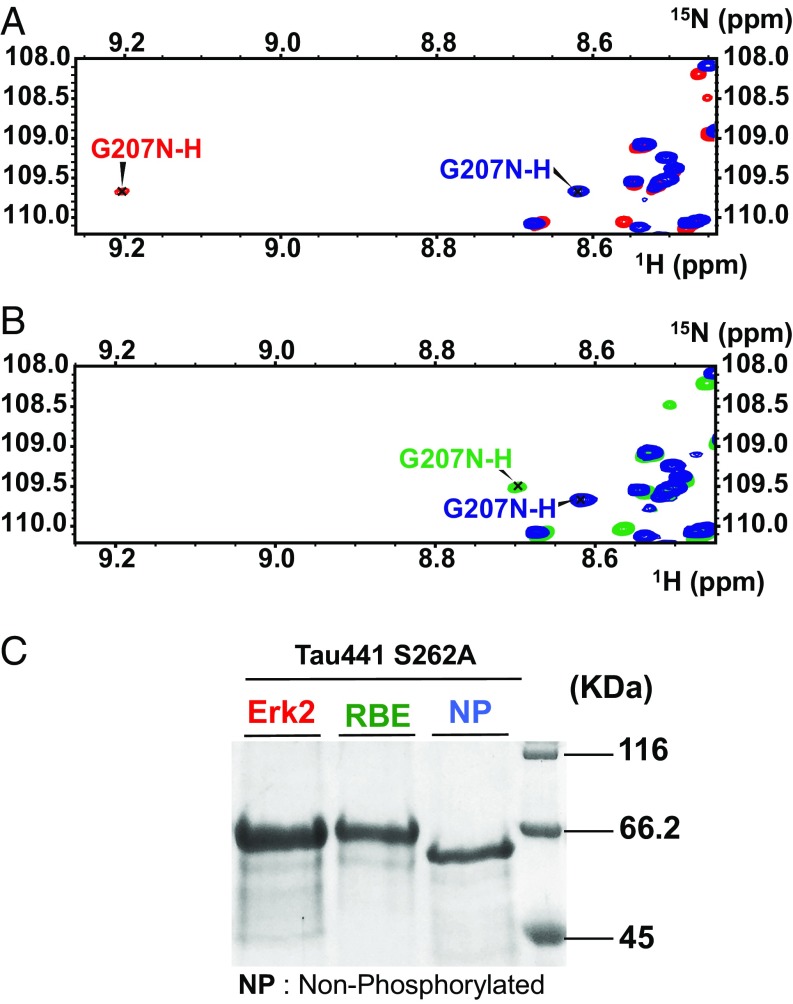
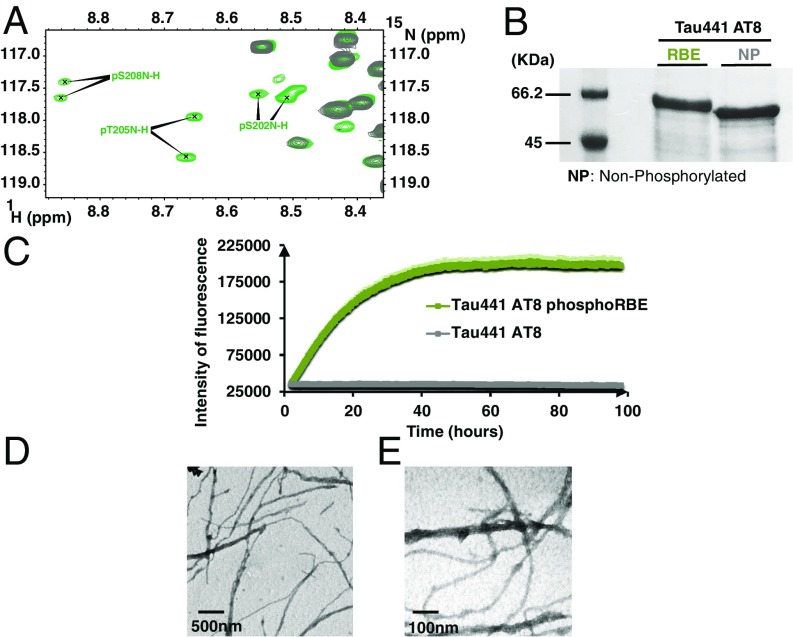
Determining the functional relationship between Tau phosphorylation and aggregation has proven a challenge owing to the multiple potential phosphorylation sites and their clustering in the Tau sequence. We use here in vitro kinase assays combined with NMR spectroscopy as an analytical tool to generate well-characterized phosphorylated Tau samples and show that the combined phosphorylation at the Ser202/Thr205/Ser208 sites, together with absence of phosphorylation at the Ser262 site, yields a Tau sample that readily forms fibers, as observed by thioflavin T fluorescence and electron microscopy. On the basis of conformational analysis of synthetic phosphorylated peptides, we show that aggregation of the samples correlates with destabilization of the turn-like structure defined by phosphorylation of Ser202/Thr205.
Keywords: Alzheimer’s disease; NMR; Tau; aggregation; phosphorylation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Goedert M, Spillantini MG. A century of Alzheimer’s disease. Science. 2006;314:777–781. - PubMed
-
- Glenner GG, Wong CW. Alzheimer’s disease: Initial report of the purification and characterization of a novel cerebrovascular amyloid protein. Biochem Biophys Res Commun. 1984;120:885–890. - PubMed
-
- Grundke-Iqbal I, et al. Microtubule-associated protein tau. A component of Alzheimer paired helical filaments. J Biol Chem. 1986;261:6084–6089. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
