

Infection-adapted emergency hematopoiesis promotes visceral leishmaniasis
- PMID: 28787450
- PMCID: PMC5560750
- DOI: 10.1371/journal.ppat.1006422
Infection-adapted emergency hematopoiesis promotes visceral leishmaniasis
Abstract
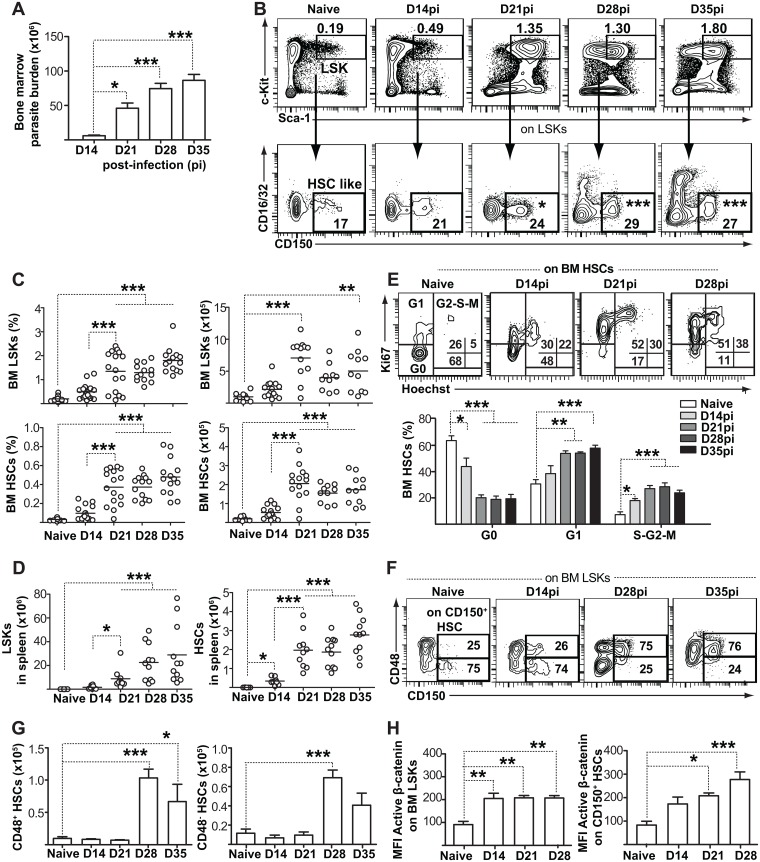
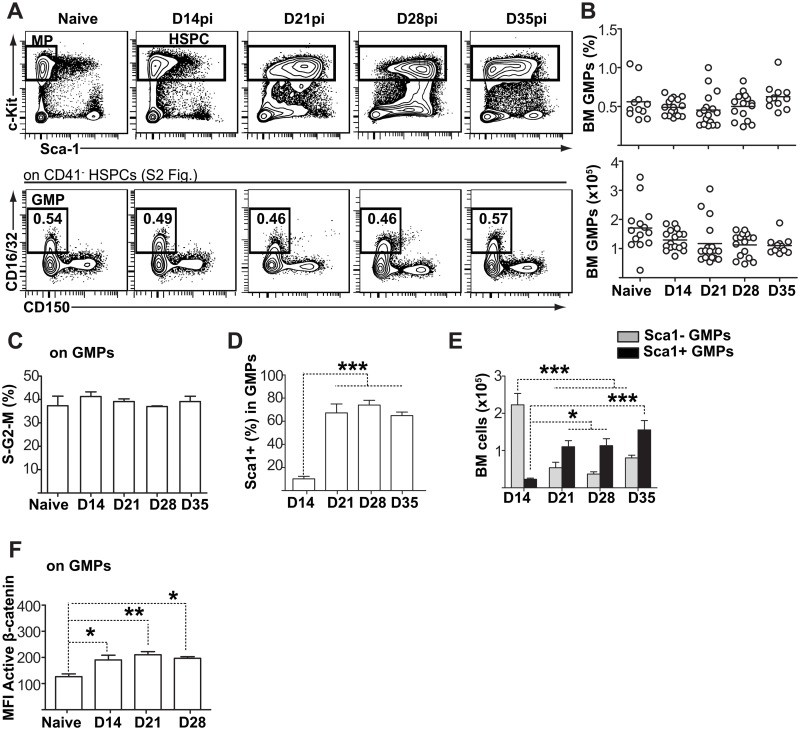
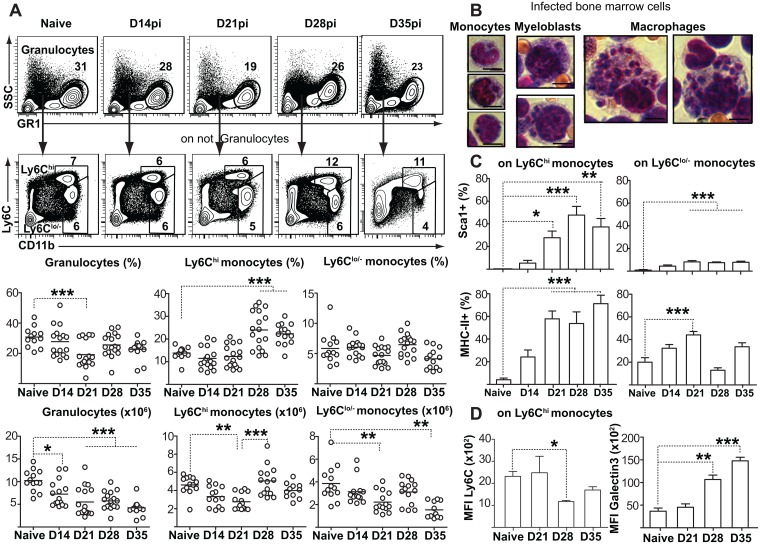
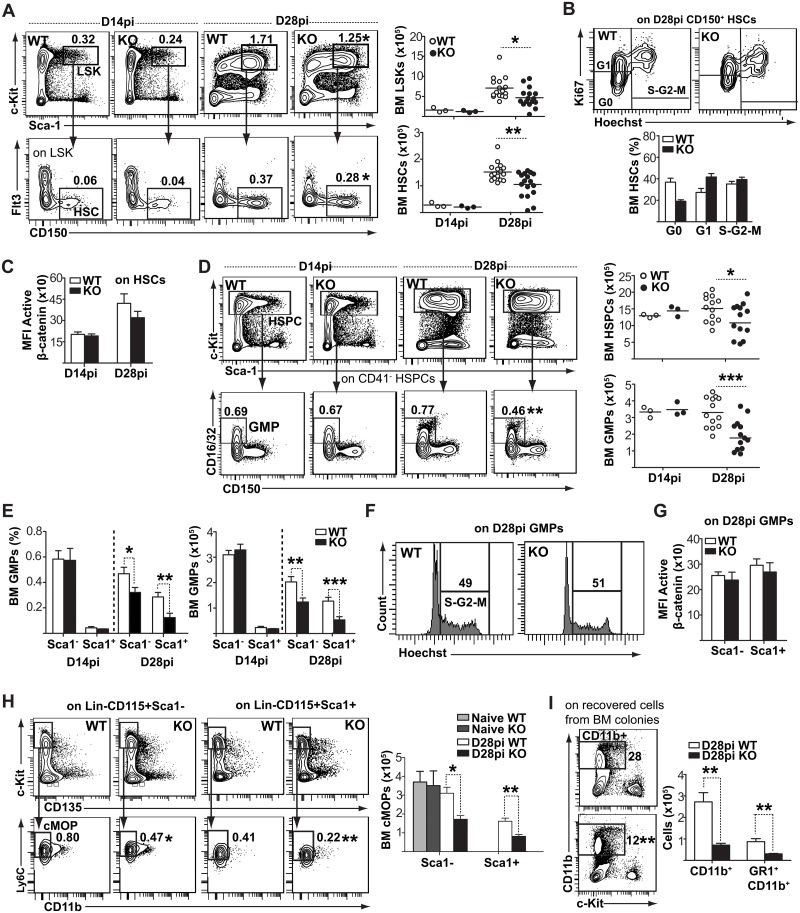
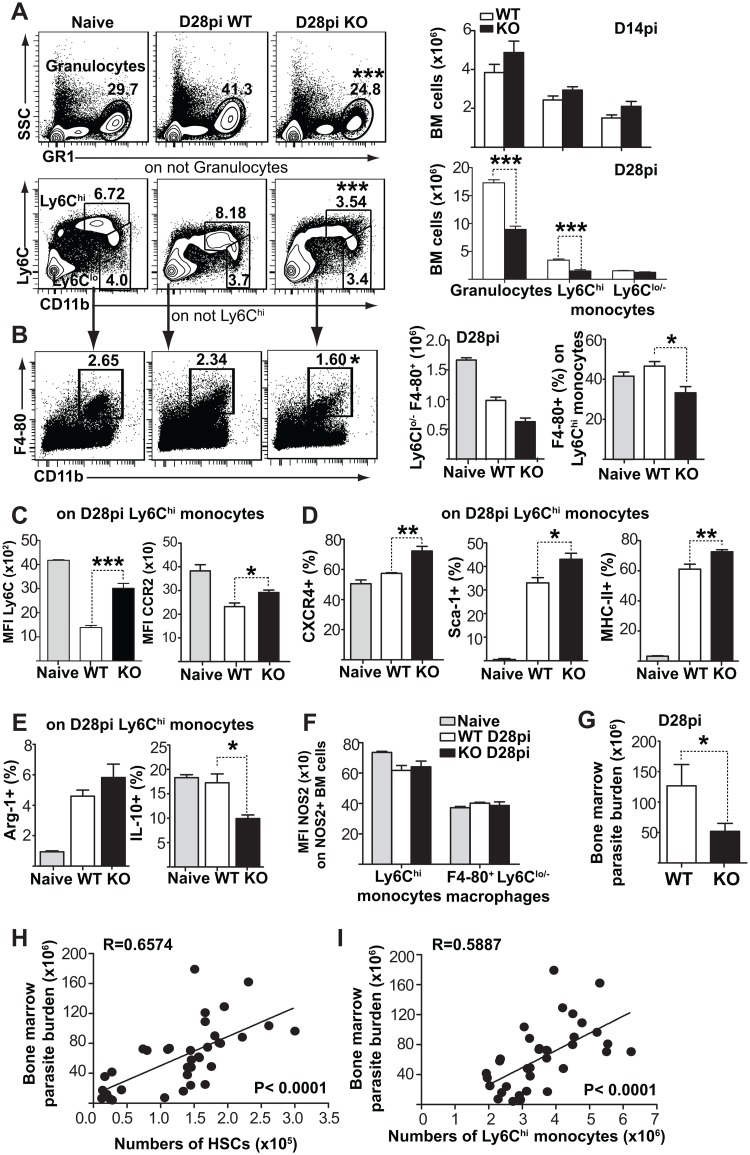
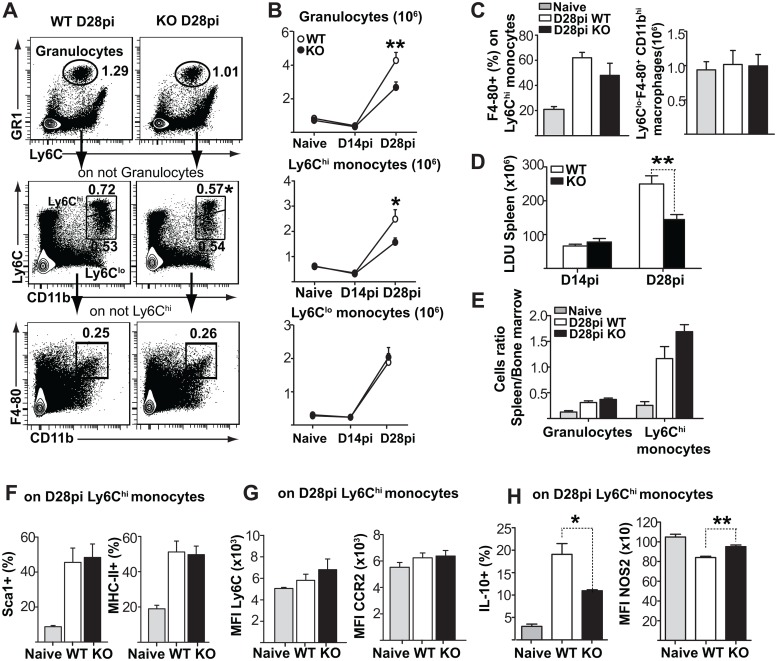
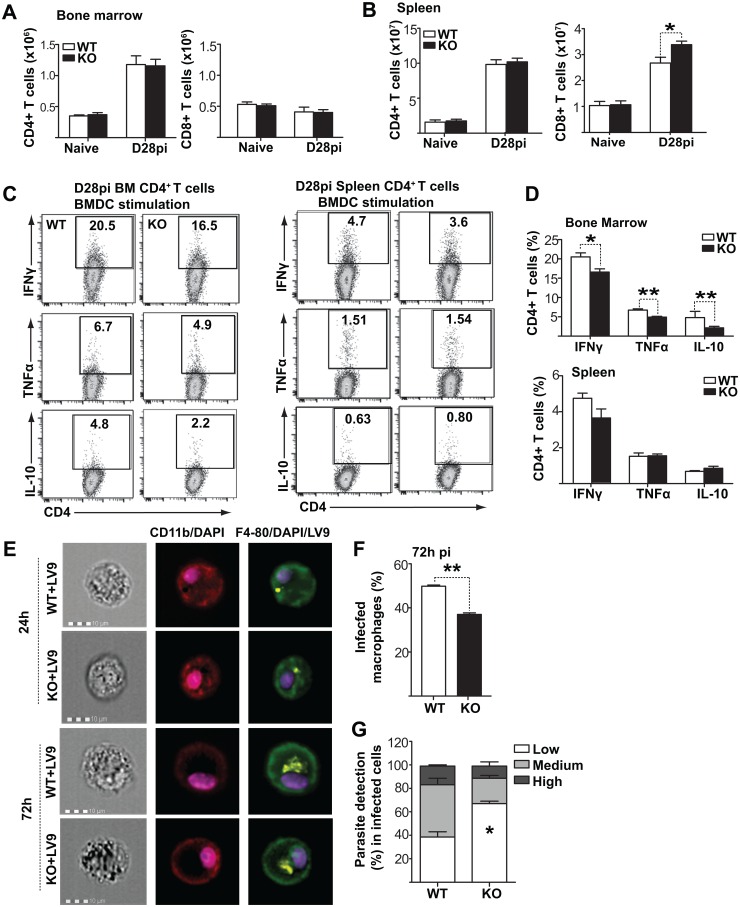
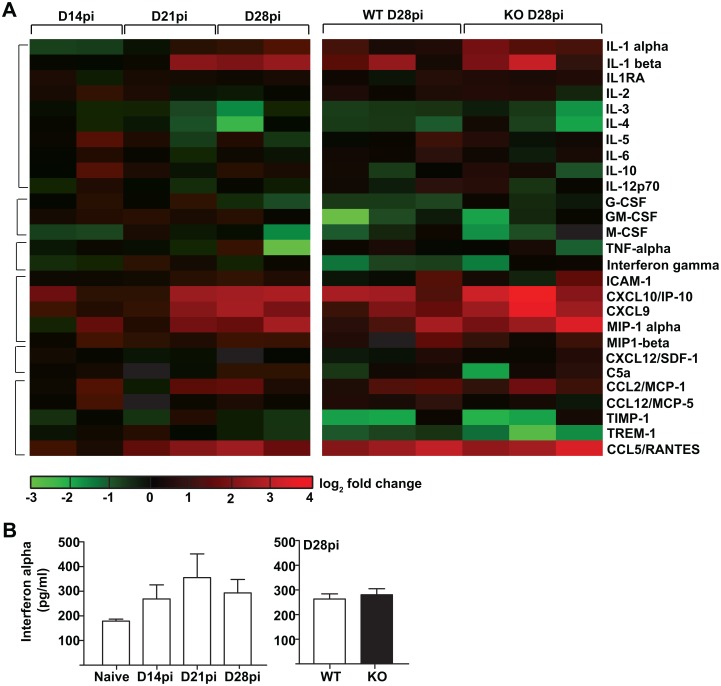
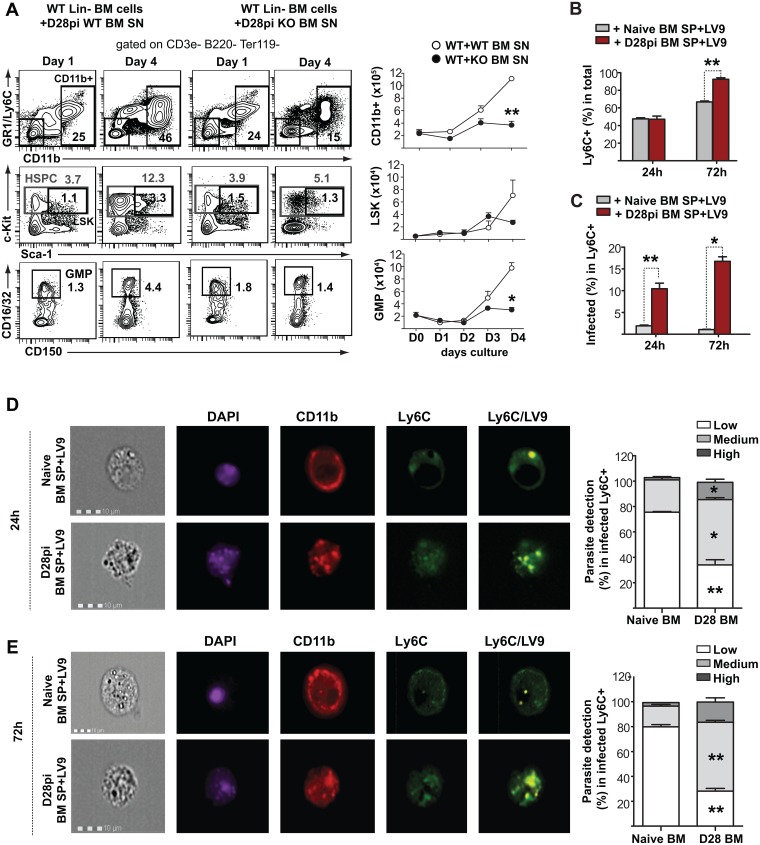
Cells of the immune system are derived from hematopoietic stem cells (HSCs) residing in the bone marrow. HSCs become activated in response to stress, such as acute infections, which adapt the bone marrow output to the needs of the immune response. However, the impact of infection-adapted HSC activation and differentiation on the persistence of chronic infections is poorly understood. We have examined here the bone marrow outcome of chronic visceral leishmaniasis and show that the parasite Leishmania donovani induces HSC expansion and skews their differentiation towards non-classical myeloid progenitors with a regulatory phenotype. Our results further suggest that emergency hematopoiesis contributes to the pathogenesis of visceral leishmaniasis, as decreased HSC expansion results in a lower parasite burden. Conversely, monocytes derived in the presence of soluble factors from the infected bone marrow environment are more permissive to infection by Leishmania. Our results demonstrate that L. donovani is able to subvert host bone marrow emergency responses to facilitate parasite persistence, and put forward hematopoiesis as a novel therapeutic target in chronic infections.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Burberry A, Zeng MY, Ding L, Wicks I, Inohara N, Morrison SJ, et al. Infection mobilizes hematopoietic stem cells through cooperative NOD-like receptor and Toll-like receptor signaling. Cell Host Microbe. 2014;15(6):779–91. doi: 10.1016/j.chom.2014.05.004 ; - DOI - PMC - PubMed
-
- Boettcher S, Gerosa RC, Radpour R, Bauer J, Ampenberger F, Heikenwalder M, et al. Endothelial cells translate pathogen signals into G-CSF-driven emergency granulopoiesis. Blood. 2014;124(9):1393–403. doi: 10.1182/blood-2014-04-570762 ; - DOI - PMC - PubMed
-
- Manz MG, Boettcher S. Emergency granulopoiesis. Nature reviews Immunology. 2014;14(5):302–14. doi: 10.1038/nri3660 . - DOI - PubMed
-
- Baldridge MT, King KY, Boles NC, Weksberg DC, Goodell MA. Quiescent haematopoietic stem cells are activated by IFN-gamma in response to chronic infection. Nature. 2010;465(7299):793–7. Epub 2010/06/11. doi: 10.1038/nature09135 ; - DOI - PMC - PubMed
-
- Griseri T, McKenzie BS, Schiering C, Powrie F. Dysregulated hematopoietic stem and progenitor cell activity promotes interleukin-23-driven chronic intestinal inflammation. Immunity. 2012;37(6):1116–29. Epub 2012/12/04. doi: 10.1016/j.immuni.2012.08.025 ; - DOI - PMC - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
