

Regulation of Osteoclast Differentiation by Myosin X
- PMID: 28790434
- PMCID: PMC5548914
- DOI: 10.1038/s41598-017-07855-9
Regulation of Osteoclast Differentiation by Myosin X
Abstract
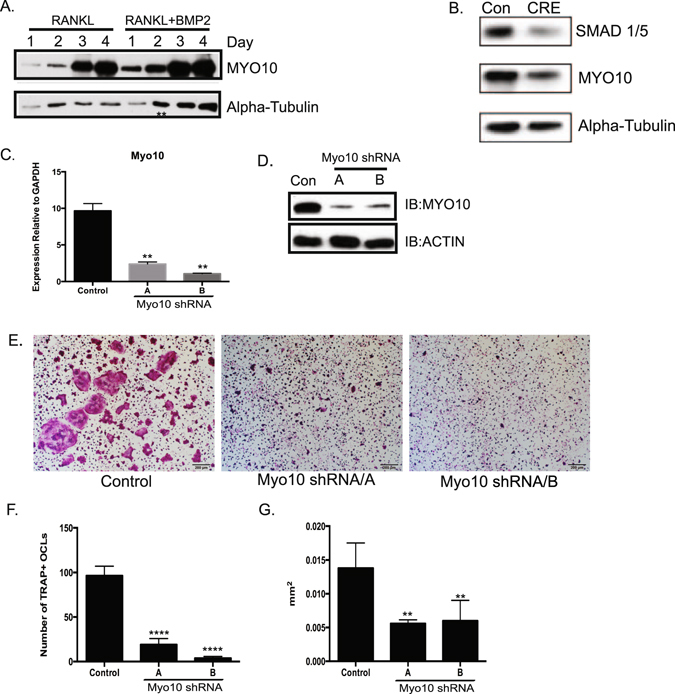
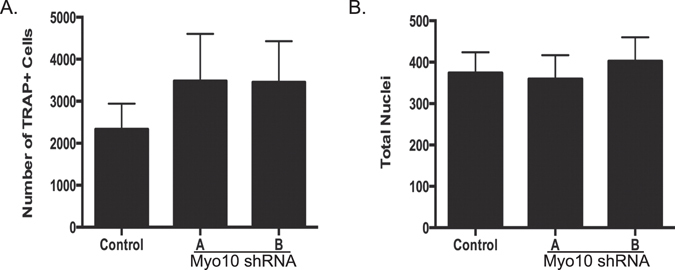
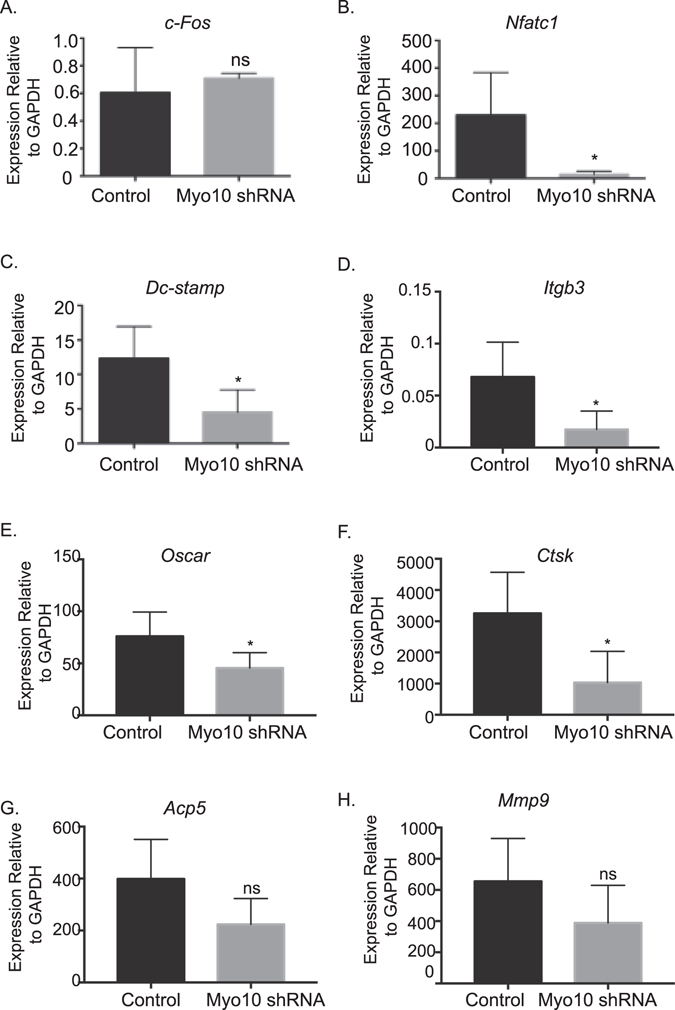
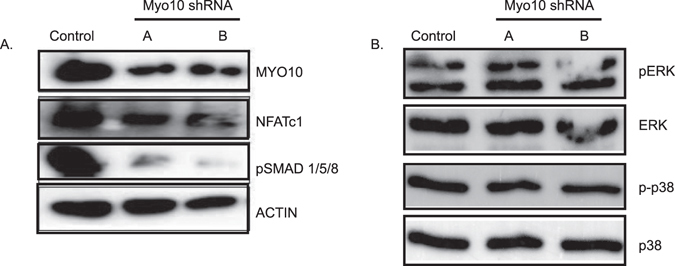
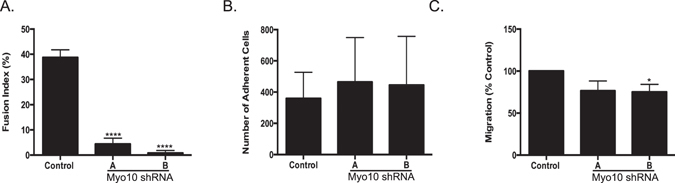
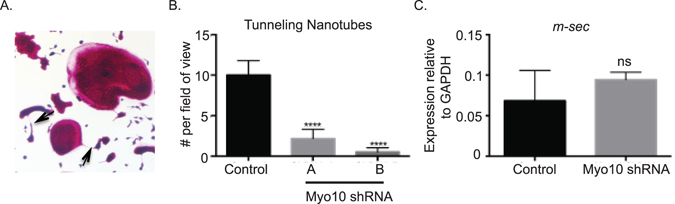
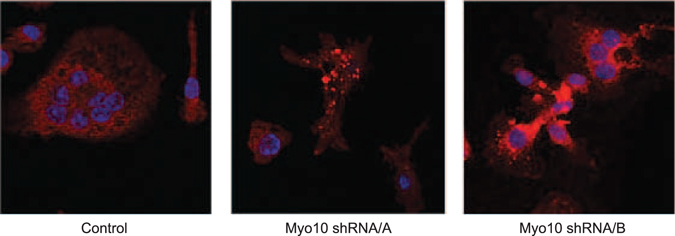
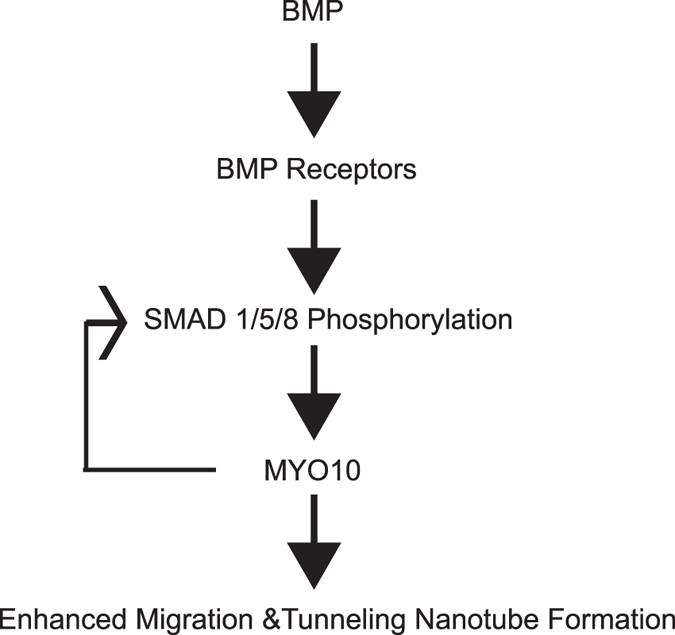
Osteoclasts begin as mononuclear cells that fuse to form multinuclear cells able to resorb bone. The mechanisms that regulate all the steps of osteoclast differentiation are not entirely known. MYO10, an unconventional myosin, has previously been shown in mature osteoclasts to play a role in attachment and podosome positioning. We determined that MYO10 is also expressed early during osteoclast differentiation. Loss of MYO10 expression in osteoclast precursors inhibits the ability of mononuclear osteoclasts to fuse into multinuclear osteoclasts. Expression of Nfatc1, Dc-stamp, Ctsk, and β 3 integrin is reduced in the osteoclasts with reduced MYO10 expression. A slight reduction in the osteoclasts ability to migrate, as well as a reduction in SMAD 1/5/8 phosphorylation are also noted with reduced MYO10 expression. Interestingly we also detected a change in the ability of the osteoclast precursors to form tunneling nanotubes (TNTs), which suggests that MYO10 may regulate the presence of TNTs through its interaction with the cytoskeletal proteins.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Vaananen, H. K. & Laitala-Leinonen, T. Osteoclast lineage and function. Arch Biochem Biophys473, 132–138, doi:S0003-9861(08)00176-8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
