

Retrovirus-Based Virus-Like Particle Immunogenicity and Its Modulation by Toll-Like Receptor Activation
- PMID: 28794025
- PMCID: PMC5640832
- DOI: 10.1128/JVI.01230-17
Retrovirus-Based Virus-Like Particle Immunogenicity and Its Modulation by Toll-Like Receptor Activation
Abstract
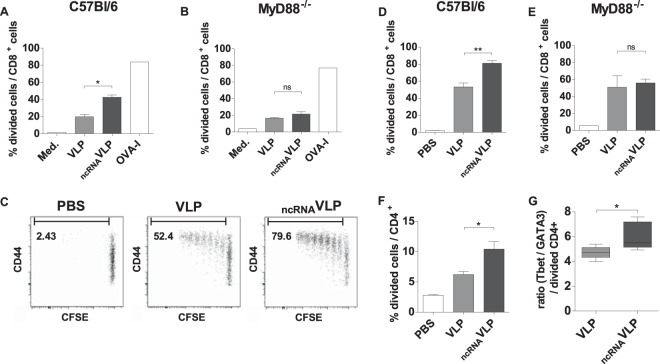
Retrovirus-derived virus-like particles (VLPs) are particularly interesting vaccine platforms, as they trigger efficient humoral and cellular immune responses and can be used to display heterologous antigens. In this study, we characterized the intrinsic immunogenicity of VLPs and investigated their possible adjuvantization by incorporation of Toll-like receptor (TLR) ligands. We designed a noncoding single-stranded RNA (ncRNA) that could be encapsidated by VLPs and induce TLR7/8 signaling. We found that VLPs efficiently induce in vitro dendritic cell activation, which can be improved by ncRNA encapsidation (ncRNAVLPs). Transcriptome studies of dendritic cells harvested from the spleens of immunized mice identified antigen presentation and immune activation as the main gene expression signatures induced by VLPs, while TLR signaling and Th1 signatures characterize ncRNAVLPs. In vivo and compared with standard VLPs, ncRNAVLPs promoted Th1 responses and improved CD8+ T cell proliferation in a MyD88-dependent manner. In an HIV vaccine mouse model, HIV-pseudotyped ncRNAVLPs elicited stronger antigen-specific cellular and humoral responses than VLPs. Altogether, our findings provide molecular evidence for a strong vaccine potential of retrovirus-derived VLPs that can be further improved by harnessing TLR-mediated immune activation.IMPORTANCE We previously reported that DNA vaccines encoding antigens displayed in/on retroviral VLPs are more efficient than standard DNA vaccines at inducing cellular and humoral immune responses. We aimed to decipher the mechanisms and investigated the VLPs' immunogenicity independently of DNA vaccination. We show that VLPs have the ability to activate antigen-presenting cells directly, thus confirming their intrinsic immunostimulatory properties and their potential to be used as an antigenic platform. Notably, this immunogenicity can be further improved and/or oriented by the incorporation into VLPs of ncRNA, which provides further TLR-mediated activation and Th1-type CD4+ and CD8+ T cell response orientation. Our results highlight the versatility of retrovirus-derived VLP design and the value of using ncRNA as an intrinsic vaccine adjuvant.
Keywords: RNA; TLR ligand; human immunodeficiency virus; immunogenicity; mice; retroviruses; vaccines; virus-like particle.
Copyright © 2017 American Society for Microbiology.
Figures




Similar articles
-
Strong CD8+ T cell antigenicity and immunogenicity of large foreign proteins incorporated in HIV-1 VLPs able to induce a Nef-dependent activation/maturation of dendritic cells.Vaccine. 2011 Apr 18;29(18):3465-75. doi: 10.1016/j.vaccine.2011.02.059. Epub 2011 Mar 5. Vaccine. 2011. PMID: 21382480
-
Incorporation of Ebola glycoprotein into HIV particles facilitates dendritic cell and macrophage targeting and enhances HIV-specific immune responses.PLoS One. 2019 May 17;14(5):e0216949. doi: 10.1371/journal.pone.0216949. eCollection 2019. PLoS One. 2019. PMID: 31100082 Free PMC article.
-
Toll-like receptor 3 adjuvant in combination with virus-like particles elicit a humoral response against HIV.Vaccine. 2016 Nov 21;34(48):5886-5894. doi: 10.1016/j.vaccine.2016.10.036. Epub 2016 Oct 27. Vaccine. 2016. PMID: 27997339 Free PMC article.
-
Bioengineered Bovine Papillomavirus L1 Protein Virus-like Particle (VLP) Vaccines for Enhanced Induction of CD8 T Cell Responses through Cross-Priming.Int J Mol Sci. 2023 Jun 7;24(12):9851. doi: 10.3390/ijms24129851. Int J Mol Sci. 2023. PMID: 37372999 Free PMC article. Review.
-
Adjuvants Enhancing Cross-Presentation by Dendritic Cells: The Key to More Effective Vaccines?Front Immunol. 2018 Dec 13;9:2874. doi: 10.3389/fimmu.2018.02874. eCollection 2018. Front Immunol. 2018. PMID: 30619259 Free PMC article. Review.
Cited by
-
All the small things: How virus-like particles and liposomes modulate allergic immune responses.Eur J Immunol. 2020 Jan;50(1):17-32. doi: 10.1002/eji.201847810. Epub 2019 Dec 15. Eur J Immunol. 2020. PMID: 31799700 Free PMC article. Review.
-
Exploiting viral sensing mediated by Toll-like receptors to design innovative vaccines.NPJ Vaccines. 2021 Oct 28;6(1):127. doi: 10.1038/s41541-021-00391-8. NPJ Vaccines. 2021. PMID: 34711839 Free PMC article. Review.
-
Viruses as tools in gene therapy, vaccine development, and cancer treatment.Arch Virol. 2022 Jun;167(6):1387-1404. doi: 10.1007/s00705-022-05432-8. Epub 2022 Apr 24. Arch Virol. 2022. PMID: 35462594 Free PMC article. Review.
-
The effects of high shear rates on the average hydrodynamic diameter measured in biomimetic HIV Gag virus-like particle dispersions.Front Bioeng Biotechnol. 2024 May 27;12:1367405. doi: 10.3389/fbioe.2024.1367405. eCollection 2024. Front Bioeng Biotechnol. 2024. PMID: 38860137 Free PMC article.
-
At-line multi-angle light scattering detector for faster process development in enveloped virus-like particle purification.J Sep Sci. 2019 Aug;42(16):2640-2649. doi: 10.1002/jssc.201900441. Epub 2019 Jun 19. J Sep Sci. 2019. PMID: 31169979 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
