

Global mRNA polarization regulates translation efficiency in the intestinal epithelium
- PMID: 28798045
- PMCID: PMC5955215
- DOI: 10.1126/science.aan2399
Global mRNA polarization regulates translation efficiency in the intestinal epithelium
Abstract
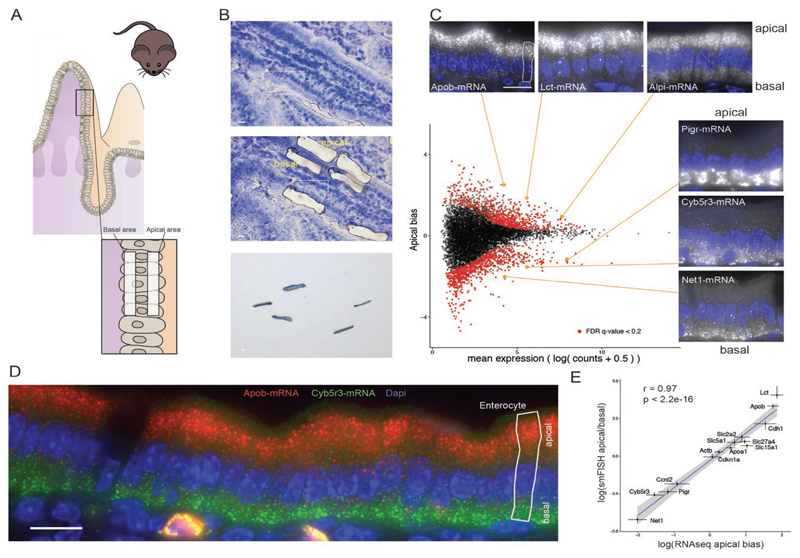
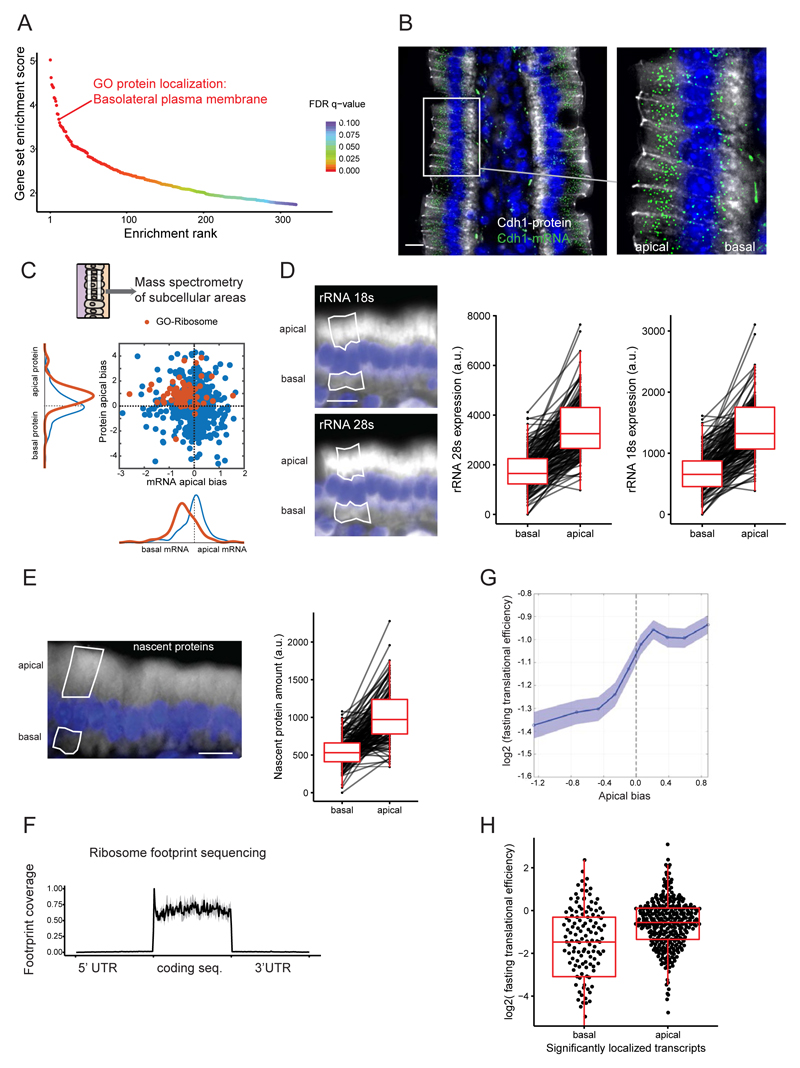
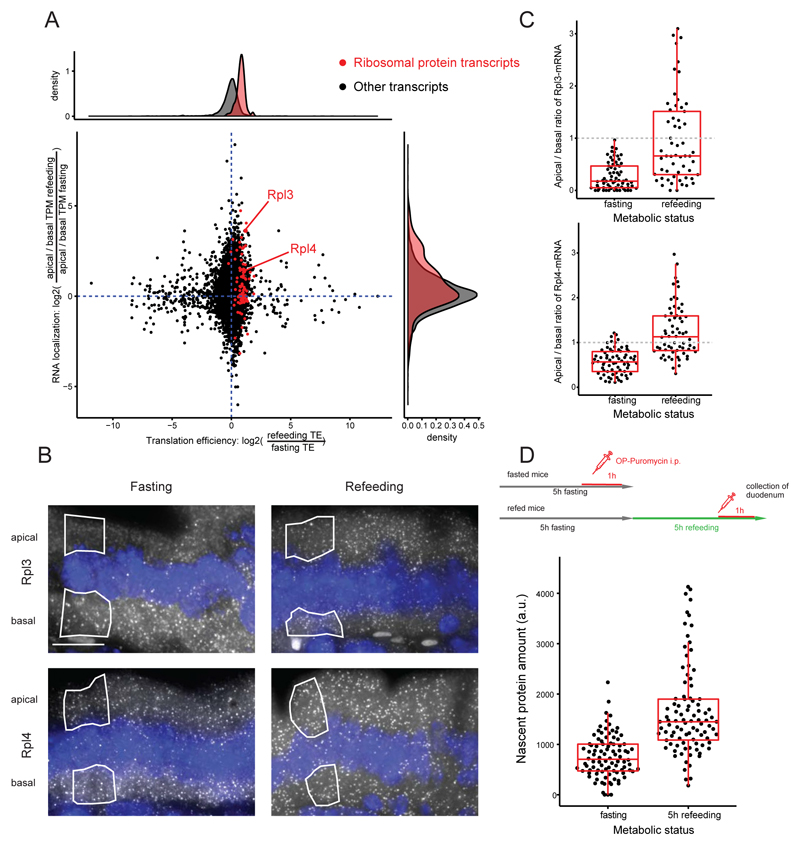
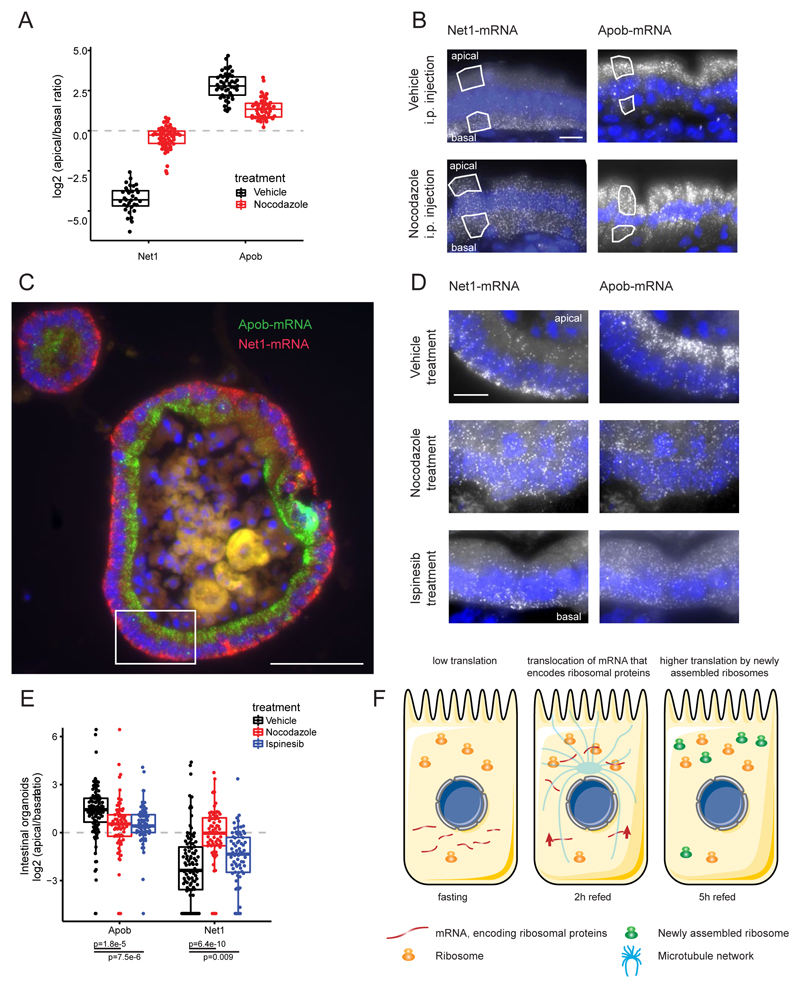
Asymmetric messenger RNA (mRNA) localization facilitates efficient translation in cells such as neurons and fibroblasts. However, the extent and importance of mRNA polarization in epithelial tissues are unclear. Here, we used single-molecule transcript imaging and subcellular transcriptomics to uncover global apical-basal intracellular polarization of mRNA in the mouse intestinal epithelium. The localization of mRNAs did not generally overlap protein localization. Instead, ribosomes were more abundant on the apical sides, and apical transcripts were consequently more efficiently translated. Refeeding of fasted mice elicited a basal-to-apical shift in polarization of mRNAs encoding ribosomal proteins, which was associated with a specific boost in their translation. This led to increased protein production, required for efficient nutrient absorption. These findings reveal a posttranscriptional regulatory mechanism involving dynamic polarization of mRNA and polarized translation.
Copyright © 2017 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
Comment in
-
mRNAs on the Move after Lunch.Dev Cell. 2017 Sep 11;42(5):439-440. doi: 10.1016/j.devcel.2017.08.017. Dev Cell. 2017. PMID: 28898674
-
RNA localization feeds translation.Science. 2017 Sep 22;357(6357):1235-1236. doi: 10.1126/science.aao5796. Science. 2017. PMID: 28935790 No abstract available.
References
-
- Lécuyer E, et al. Global analysis of mrna localization reveals a prominent role in organizing cellular architecture and function. Cell. 2007;131:174–187. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
