

MicroRNA-132 promotes fibroblast migration via regulating RAS p21 protein activator 1 in skin wound healing
- PMID: 28798331
- PMCID: PMC5552762
- DOI: 10.1038/s41598-017-07513-0
MicroRNA-132 promotes fibroblast migration via regulating RAS p21 protein activator 1 in skin wound healing
Abstract
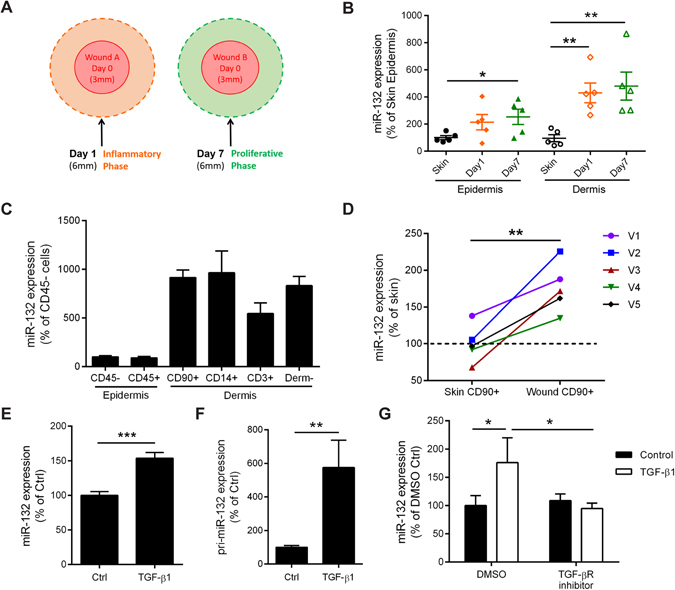
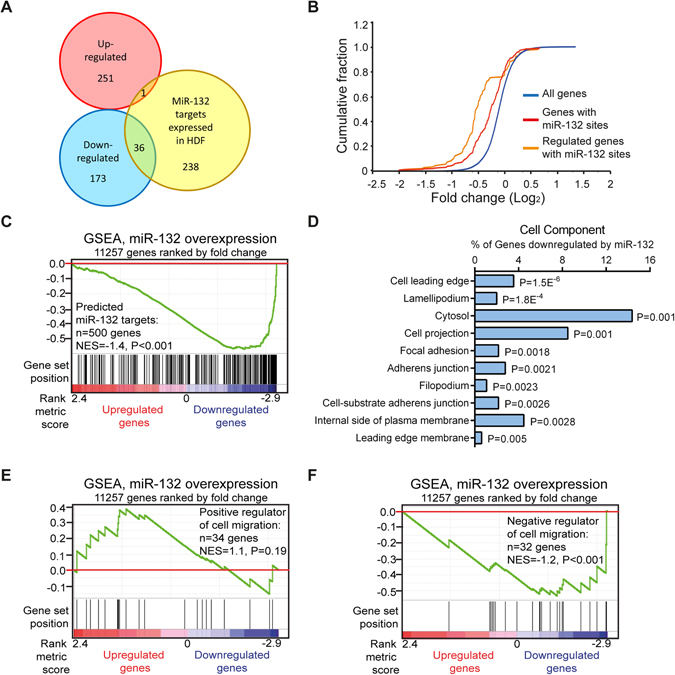
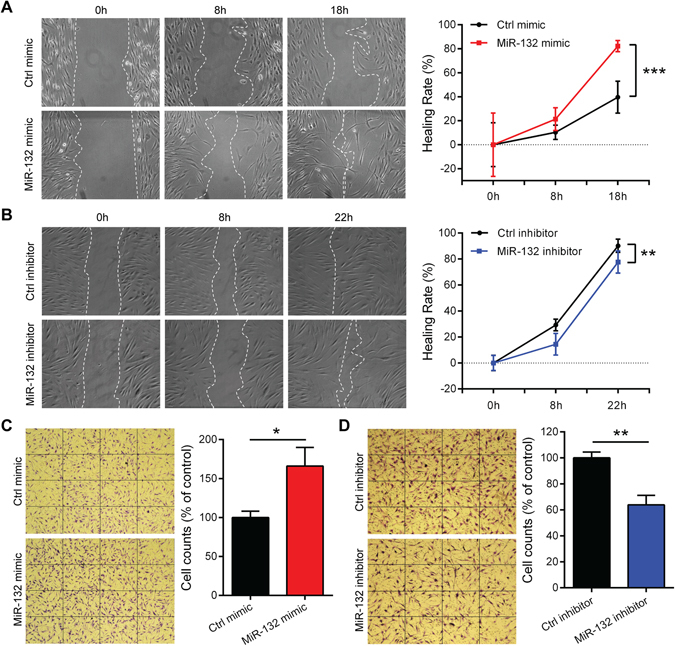
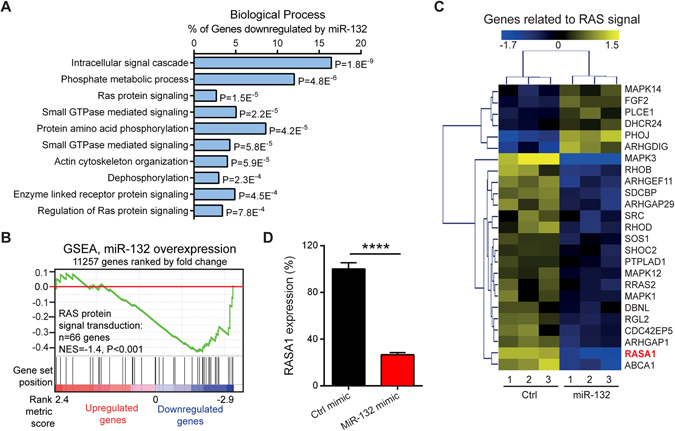
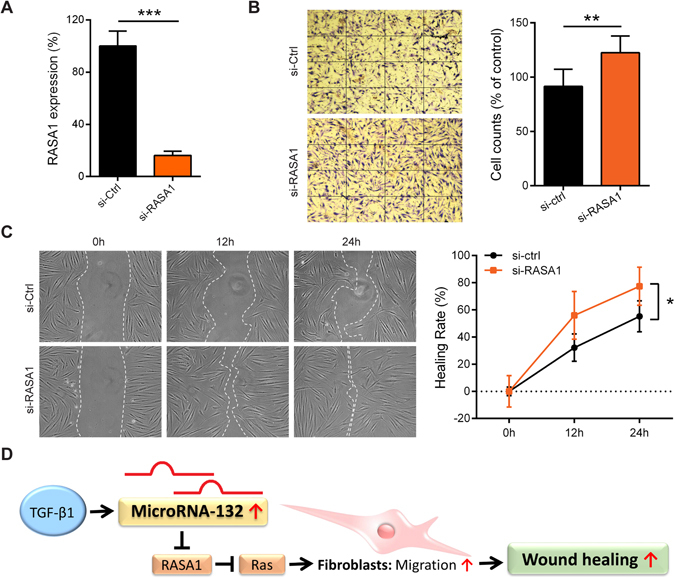
MicroRNA (miR)-132 has been identified as a top up-regulated miRNA during skin wound healing and its inhibition impairs wound repair. In a human in vivo surgical wound model, we showed that miR-132 was induced in epidermal as well as in dermal wound-edge compartments during healing. Moreover, in a panel of cells isolated from human skin wounds, miR-132 was found highly expressed in human dermal fibroblasts (HDFs). In HDFs, miR-132 expression was upregulated by TGF-β1. By overexpression or inhibition of miR-132, we showed that miR-132 promoted HDF migration. Mechanistically, global transcriptome analysis revealed that RAS signaling pathway was regulated by miR-132 in HDFs. We found that RAS p21 protein activator 1 (RASA1), a known target of miR-132, was downregulated in HDFs upon miR-132 overexpression. Silencing of RASA1 phenocopied the pro-migratory effect of miR-132. Collectively, our study reveals an important role for miR-132 in HDFs during wound healing and indicates a therapeutic potential of miR-132 in hard-to-heal skin wounds.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
