

Review
doi: 10.2174/0929866524666170811125109.
Glycan Phosphorylases in Multi-Enzyme Synthetic Processes
Affiliations
- PMID: 28799504
- PMCID: PMC5688430
- DOI: 10.2174/0929866524666170811125109
Item in Clipboard
Review
Glycan Phosphorylases in Multi-Enzyme Synthetic Processes
Protein Pept Lett.
2017.
Abstract
Glycoside phosphorylases catalyse the reversible synthesis of glycosidic bonds by glycosylation with concomitant release of inorganic phosphate. The equilibrium position of such reactions can render them of limited synthetic utility, unless coupled with a secondary enzymatic step where the reaction lies heavily in favour of product. This article surveys recent works on the combined use of glycan phosphorylases with other enzymes to achieve synthetically useful processes.
Keywords: Phosphorylase; biofuel; cellodextrin; disaccharide; high-value products; α-glucan.
Copyright© Bentham Science Publishers; For any queries, please email at epub@benthamscience.org.
Figures
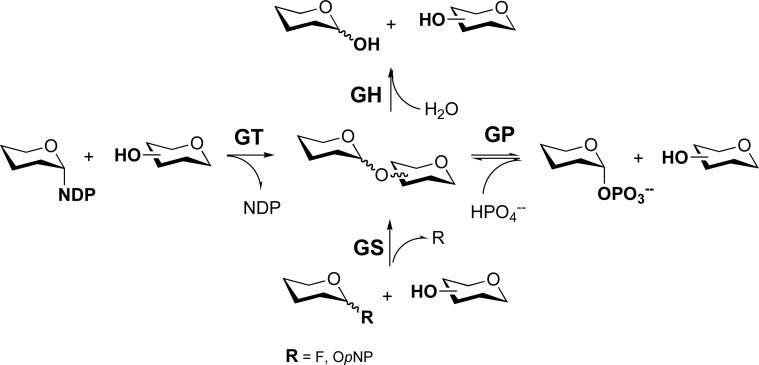
Enzymes involved in glycosidic bond formation and cleavage. GT = glycosyltransferase; GP = glycoside phosphorylase; GH = glycoside hydrolase; GS = glycoside synthase; NDP = nucleotide diphosphate; OpNP = O-para-nitrophenyl.
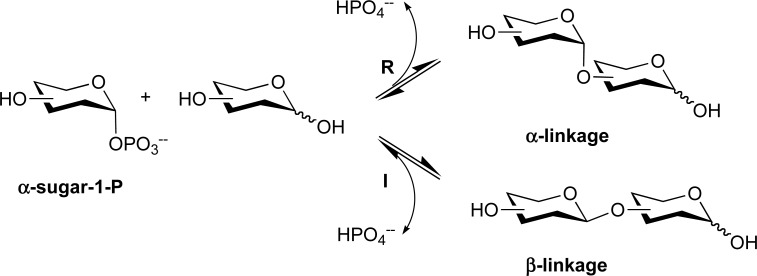
Reaction mechanism of glycoside phosphorylases. R, retaining; I, inverting.
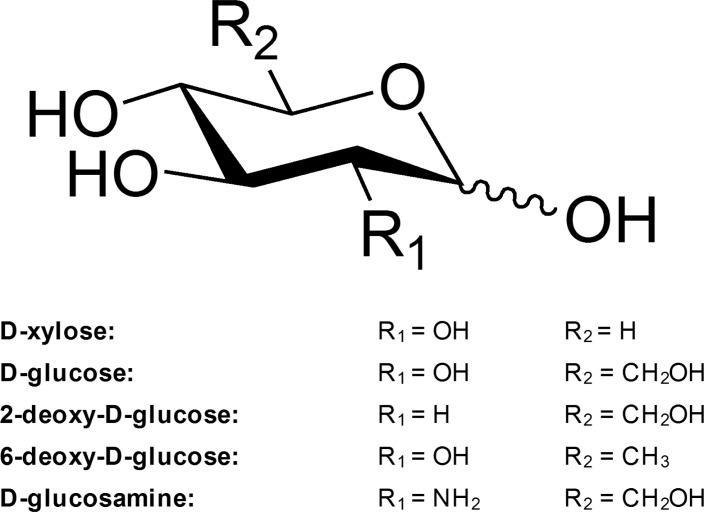
Structure illustration for CBP acceptors [25]. R1 and R2, which are linked to C-2 and C-6 respectively, are substituted by different functional groups for each of the aforementioned acceptors.
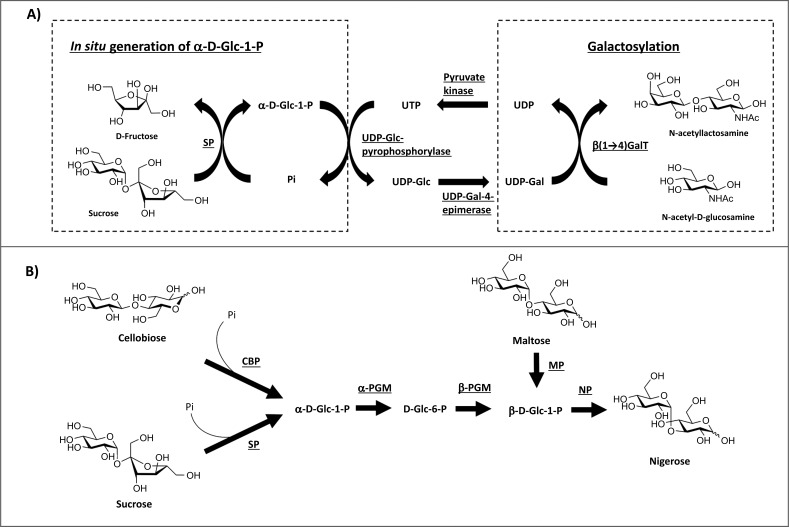
A) In situ generation of α-D-Glc-1-phosphate by SP coupling with galactosylation of N-acetylglucosamine [27]. β-(1→4)-GalT = β-(1→4)-galactosyltransferase; UDP = uridine diphosphate; UTP = uridine triphosphate; Pi = inorganic phosphate; SP = sucrose phosphorylase. B) Enzymatic production of nigerose from 4 different starting materials (cellobiose, starch, sucrose and maltose) [28]. Cellobiose, starch and sucrose were used for the production of α-D-Glc-1-phosphate, which was converted to β-anomer, and subsequently used by nigerose phosphorylase for nigerose production. Maltose was also phosphorolysed by maltose phosphorylase to generate β-D-Glc-1-phophate which can be used directly by nigerose phosphorylase. Pi = inorganic phosphate; CBP = cellobiose phosphorylase; SP = sucrose phosphorylase; MP = maltose phosphorylase; NP = nigerose phosphorylase; PGM = phosphoglucomutase.
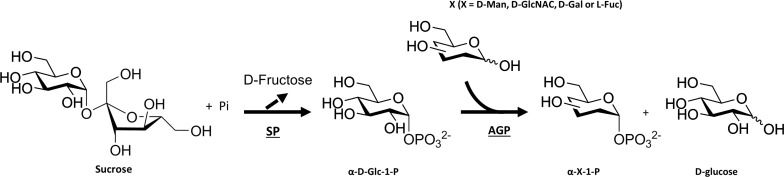
Coupling SP with AGP for the enzymatic production of a variety of sugar-1-phosphate with axial selectivity at the anomeric position [29]. Sucrose was used by SP to generate α-D-Glc-1-phosphate, which was subsequently or simultaneously used in combination with 4 different acceptors, namely D-Man, D-GlcNAc, D-Gal or L-Fuc, by AGP for the production of the corresponding sugar-1-phosphates. SP = sucrose phosphorylase and AGP = α-glucose-1-phosphatase.
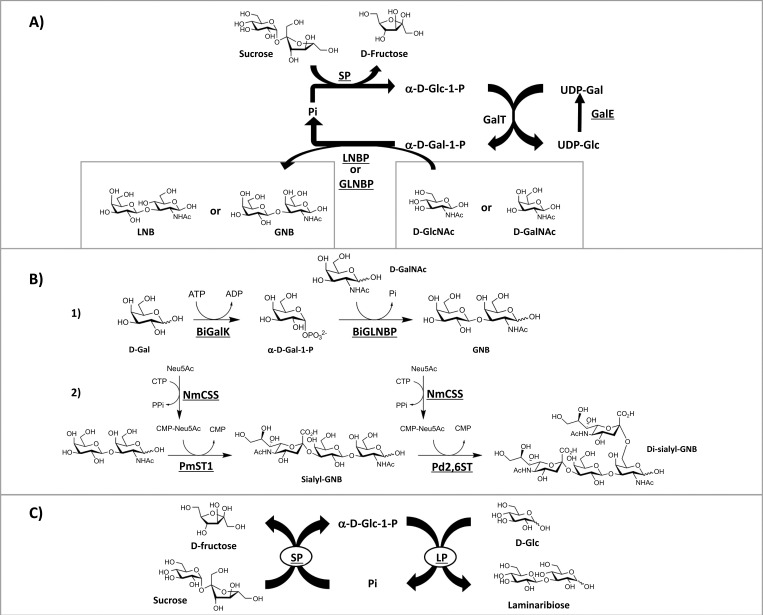
A) Two biosynthetic routes for GNB and LNB production proposed by Nishimoto and Kitaoka [35, 40]. For lacto-N-biose (LNB) production, LNBP and D-GlcNAc were used as the final catalyst and substrate respectively. LNBP and D-GlcNAc are substituted by GLNBP and D-GalNAc, respectively, for the production of GNB. B) Enzymatic production of GNB and sialylated-GNB proposed by Li and co-workers [41]. (1) BiGalK was used to convert D-Gal to α-D-Gal-1-P, which was utilised as a sugar donor by BiGLNBP with D-GalNAc as an acceptor to produce GNB. (2) Sialyl-GNB was formed by transferring Neu5Ac from CMP-Neu5Ac onto D-Gal of GNB via α-(2→3) linkage by the activity of PmST1. The sialyl-GNB was further sialylated at C-6 of D-GalNAc by Pd2,6ST via α-(2→6) linkage to form disialyl-GNB. BiGLNBP = GNB/LNB phosphorylase from Bifidobacterium infantis; BiGalK = galactokinase from Bifidobacterium infantis; NmCSS = CMP-Sia synthetase from Neisseria meningitides; PmST1 = α-(2→3)-sialyltransferase 1 from Pasteurella multocida; Pd2,6ST = α-(2→6)-sialyltransferase from Photobacerium damselae.
C) Combination of immobilised LP from Euglena gracilis and SP can be used for the production of laminaribiose using sucrose as a starting material [42]. SP = sucrose phosphorylase; Pi = inorganic phosphate; LP = laminaribiose phosphorylase.
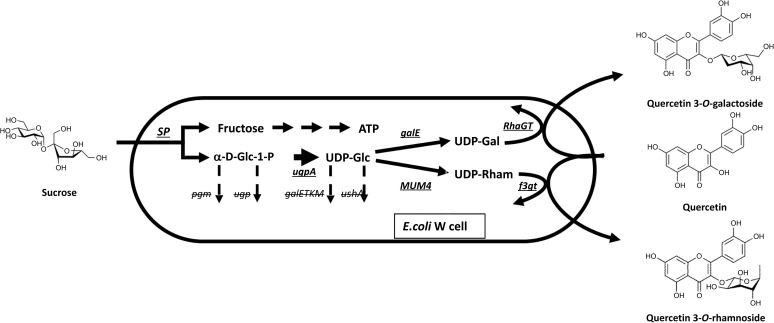
Metabolic engineering of E. coli for in vivo glycosylation platform of a flavonol molecule [49]. Underlined genes were introduced into E. coli for overexpression of the enzymes that are essential for production of desirable sugar metabolites, whereas erased genes were deleted from E. coli genome to prevent the breakdown of α-D-Glc-1-phosphate and UPD-Glc.
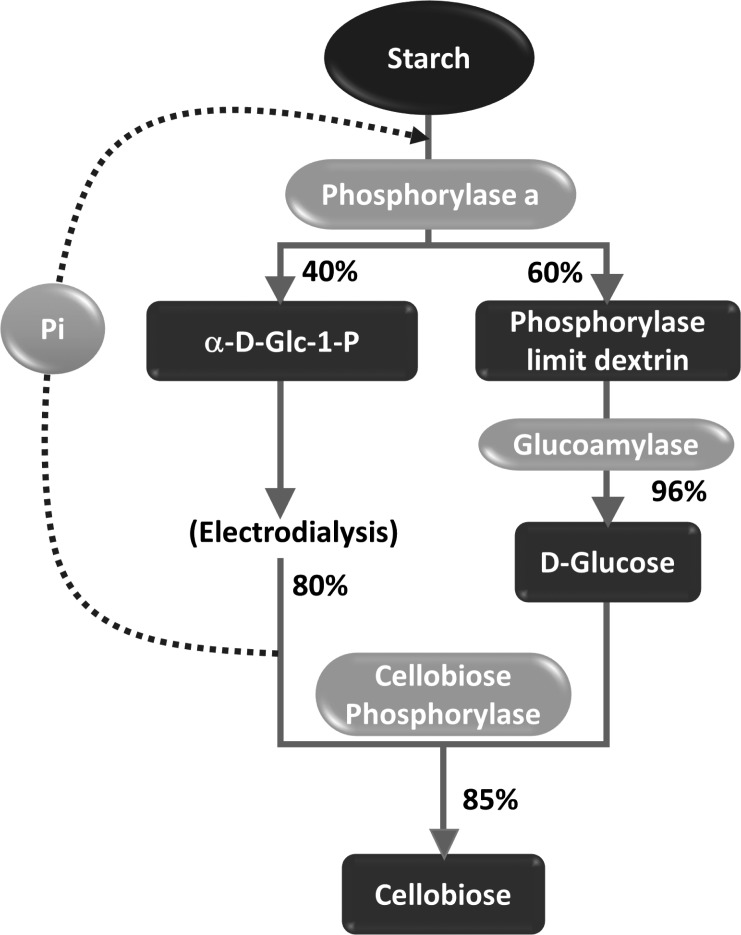
Reaction scheme depicting the conversion of starch to cellobiose using a combination of α-glucan phosphorylase and cellobiose phosphorylase [54]. Pi = inorganic phosphate.
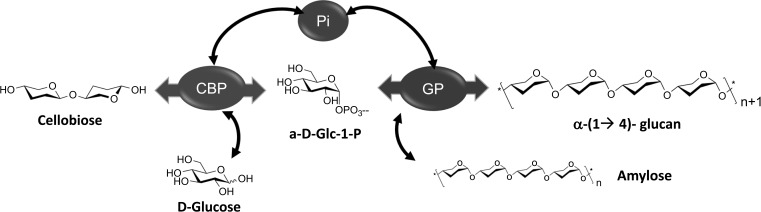
Reaction scheme depicting the combination of cellobiose phosphorylase (CBP) and α-glucan phosphorylase (GP) for the synthesis of synthetic amylose [53]. Pi = inorganic phosphate.
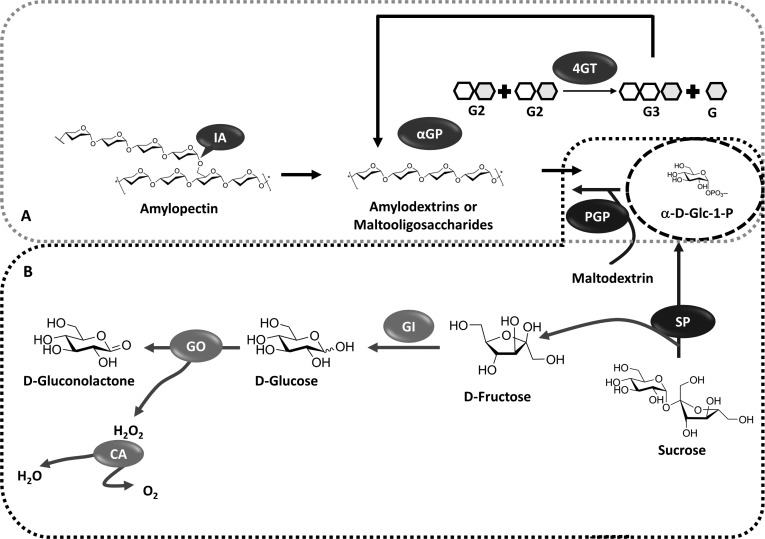
Multi-enzyme one-pot reaction schemes for generation of α-D-Glc-1-phosphate. A) Enzymatic synthesis of α-D-Glc-1-phosphate from corn starch [55]. α-D-Glc-1-phosphate synthesis corn starch-mediated by thermostable α-glucan phosphorylase (αGP) from Thermotoga maritima. The hyper-thermostable isoamylase (IA) from Sulfolobus tokodaii was added for enhancing the α-D-Glc-1-phosphate yield. A third enzyme, 4-glucanotransferase (4GT) from Thermococcus litoralis was added to the reaction mixture for increasing the α-D-Glc-1-phosphate titre through maltose and maltotriose utilisation. B) One-pot scheme for enzymatic synthesis of α-D-Glc-1-phosphate from sucrose [56]. SP = sucrose phosphorylase from Thermoanaerobacterium thermosaccharolyticum (EC 2.4.1.7); PGP = potato α-glucan phosphorylase (EC 2.4.1.1); GI = glucose isomerase (EC 5.3.1.5); GO = glucose oxidase (EC 1.1.3.4); CA = catalase (EC 1.11.1.6).
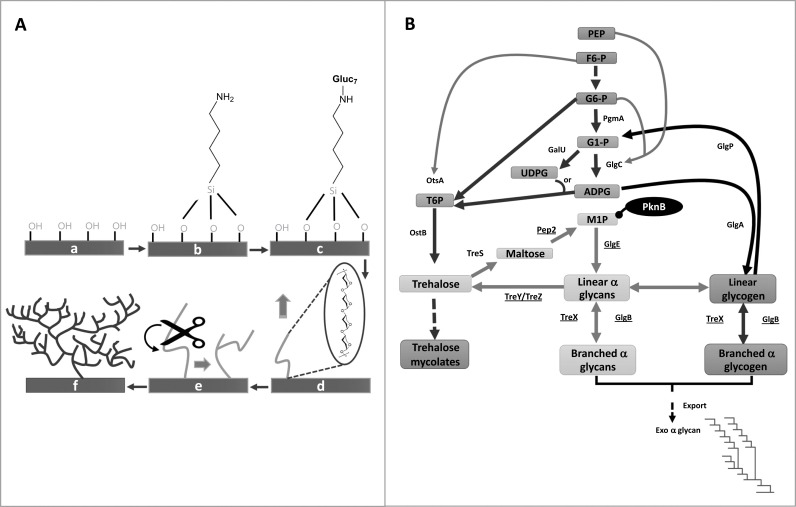
A) Combined use of potato α-glucan phosphorylase (αGP) and glycogen branching enzyme (GBE) from Deinococcus geothermalis for the generation of hyperbranched polysaccharides on silicon wafer [59]. (a) Oxidized Si wafer; (b) aminosilanized wafer; (c) maltoheptaose functionalised surface; (d) enzyme-catalysed growth of linear chain by αGP; (e) catalytic action of GBE; (f) resulting hyperbranched polysaccharide after the combined biocatalysis. B) Metabolic pathways associated with α-glucan and glycogen metabolism in Mycobacterium tuberculosis [61, 63]. The genes involved in the GlgE pathway are underlined. The pathway is negatively regulated by the serine/threonine kinase, PknB. The pathway has been exploited for generation of branched glycans using a combination of the GlgE and GlgB enzymes.
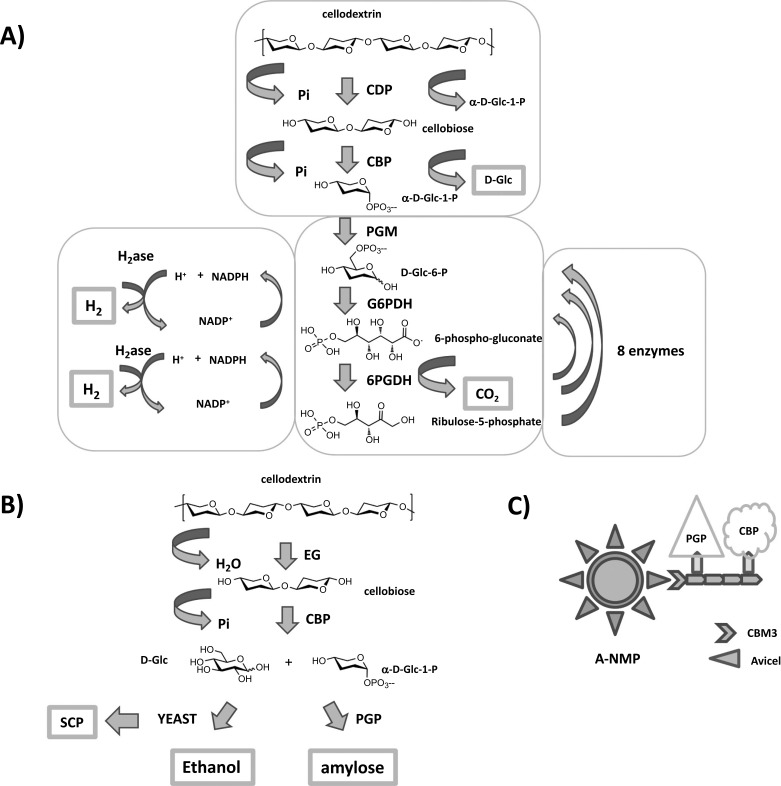
A) Production of H2 from enzymatic degradation of cellulosic biomass [78]. Reaction products are in bold boxes. Pi = inorganic phosphate; CDP = cellodextrin phosphorylase; CBP = cellobiose phosphorylase; PGM = phosphoglucomutase; G6PDH = Glc-6-phosphate dehydrogenase; 6PGDH = 6-phosphogluconate dehydrogenase; NADP+ = nicotinamide adenine dinucleotide phosphate (oxidised form); NADPH = nicotinamide adenine dinucleotide phosphate (reduced form). B) Enzymatic degradation of cellulosic waste into valuable products [79]. Cellulose enzymatic hydrolysis leads to the production of ethanol by yeast fermentation, amylose by potato a-glucan phosphorylase (PGP), and single cell protein (SCP). All products are in bold boxes. EG = endo-glucanase; Pi = inorganic phosphate; CBP = cellobiose phosphorylase. C) Co-immobilisation of CBM3, PGP and CBP onto Avicel-nano magnetic particles (A-NMPs). CBM3 = carbohydrate binding module 3.
Similar articles
-
Characterization of Ruminococcus albus cellodextrin phosphorylase and identification of a key phenylalanine residue for acceptor specificity and affinity to the phosphate group.FEBS J. 2013 Sep;280(18):4463-73. doi: 10.1111/febs.12408. Epub 2013 Jul 19. FEBS J. 2013. PMID: 23802549
-
Crystal Structure and Substrate Recognition of Cellobionic Acid Phosphorylase, Which Plays a Key Role in Oxidative Cellulose Degradation by Microbes.J Biol Chem. 2015 Jul 24;290(30):18281-92. doi: 10.1074/jbc.M115.664664. Epub 2015 Jun 3. J Biol Chem. 2015. PMID: 26041776 Free PMC article.
-
Recombinant production and biochemical characterization of a hyperthermostable alpha-glucan/maltodextrin phosphorylase from Pyrococcus furiosus.Archaea. 2008 Dec;2(3):169-76. doi: 10.1155/2008/549759. Archaea. 2008. PMID: 19054743 Free PMC article.
-
Enzymatic synthesis using glycoside phosphorylases.Carbohydr Res. 2015 Feb 11;403:23-37. doi: 10.1016/j.carres.2014.06.010. Epub 2014 Jun 18. Carbohydr Res. 2015. PMID: 25060838 Free PMC article. Review.
-
Carbohydrate synthesis by disaccharide phosphorylases: reactions, catalytic mechanisms and application in the glycosciences.Biotechnol J. 2010 Dec;5(12):1324-38. doi: 10.1002/biot.201000217. Biotechnol J. 2010. PMID: 21154671 Review.
Cited by
-
Recent advances in enzymatic synthesis of β-glucan and cellulose.Carbohydr Res. 2021 Oct;508:108411. doi: 10.1016/j.carres.2021.108411. Epub 2021 Jul 24. Carbohydr Res. 2021. PMID: 34392134 Free PMC article. Review.
-
Engineering cascade biocatalysis in whole cells for bottom-up synthesis of cello-oligosaccharides: flux control over three enzymatic steps enables soluble production.Microb Cell Fact. 2022 Apr 9;21(1):61. doi: 10.1186/s12934-022-01781-w. Microb Cell Fact. 2022. PMID: 35397553 Free PMC article.
-
Recent progress in synthesis of carbohydrates with sugar nucleotide-dependent glycosyltransferases.Curr Opin Chem Biol. 2021 Apr;61:81-95. doi: 10.1016/j.cbpa.2020.10.007. Epub 2020 Dec 10. Curr Opin Chem Biol. 2021. PMID: 33310623 Free PMC article. Review.
-
The structure of a GH149 β-(1 → 3) glucan phosphorylase reveals a new surface oligosaccharide binding site and additional domains that are absent in the disaccharide-specific GH94 glucose-β-(1 → 3)-glucose (laminaribiose) phosphorylase.Proteins. 2019 Oct;87(10):885-892. doi: 10.1002/prot.25745. Epub 2019 Jun 6. Proteins. 2019. PMID: 31134667 Free PMC article.
-
Preparative and Kinetic Analysis of β-1,4- and β-1,3-Glucan Phosphorylases Informs Access to Human Milk Oligosaccharide Fragments and Analogues Thereof.Chembiochem. 2020 Apr 1;21(7):1043-1049. doi: 10.1002/cbic.201900440. Epub 2019 Dec 30. Chembiochem. 2020. PMID: 31657512 Free PMC article.
References
-
- Field R.A. Glycobiology: Challenging reaction equilibria. Nat. Chem. Biol. 2011;7:658–659. - PubMed
-
- Puchart V. Glycoside phosphorylases: Structure, catalytic properties and biotechnological potential. Biotechnol. Adv. 2015;33:261–276. - PubMed
-
- Nakai H., Kitaoka M., Svensson B., Ohtsubo K.I. Recent development of phosphorylases possessing large potential for oligosaccharide synthesis. Curr. Opin. Chem. Biol. 2013;17:301–309. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
