

Fungal genome and mating system transitions facilitated by chromosomal translocations involving intercentromeric recombination
- PMID: 28800596
- PMCID: PMC5568439
- DOI: 10.1371/journal.pbio.2002527
Fungal genome and mating system transitions facilitated by chromosomal translocations involving intercentromeric recombination
Abstract
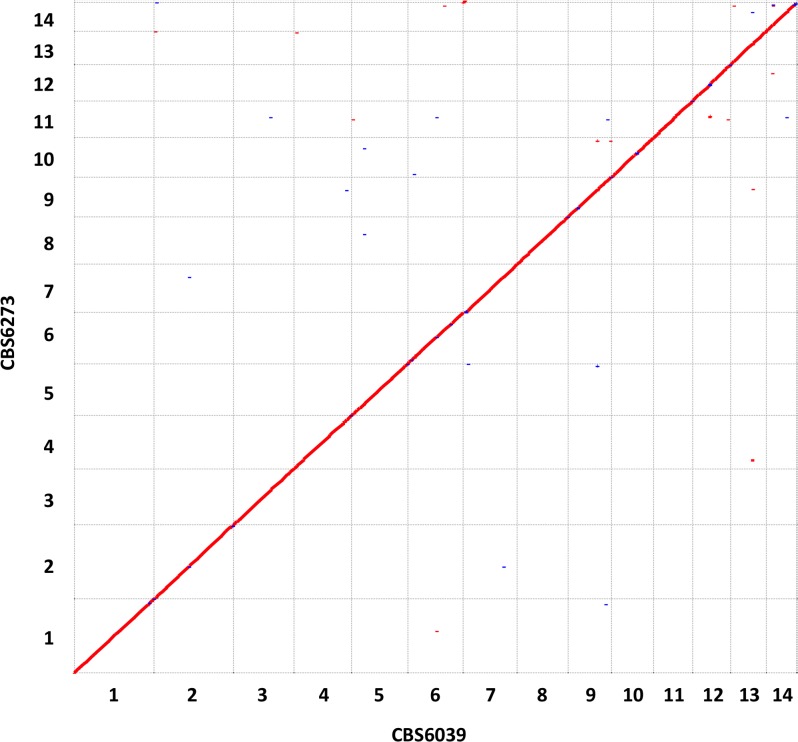
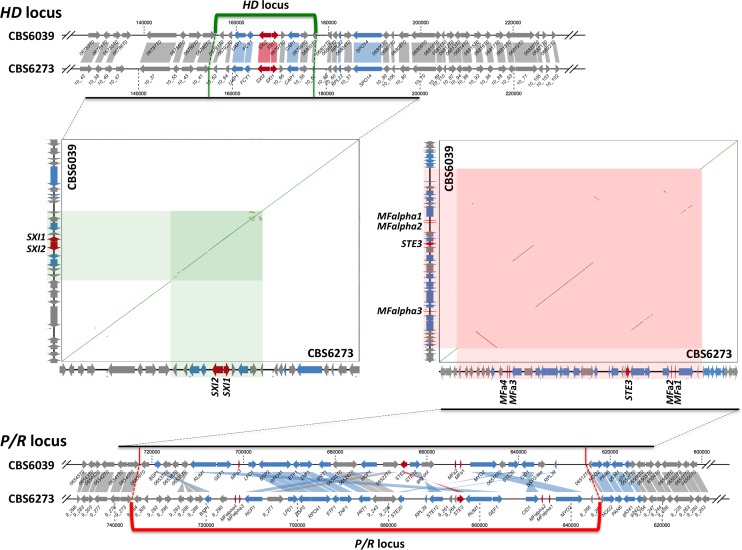
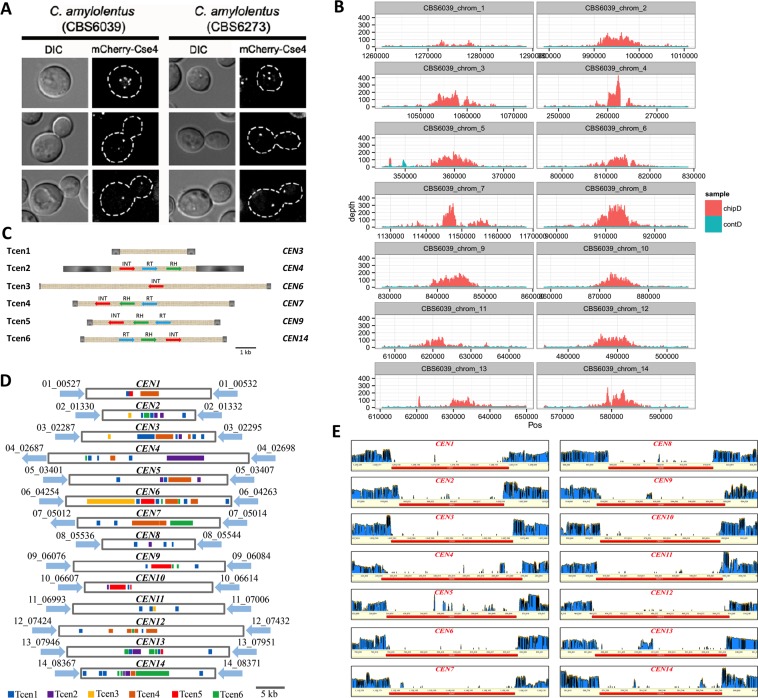
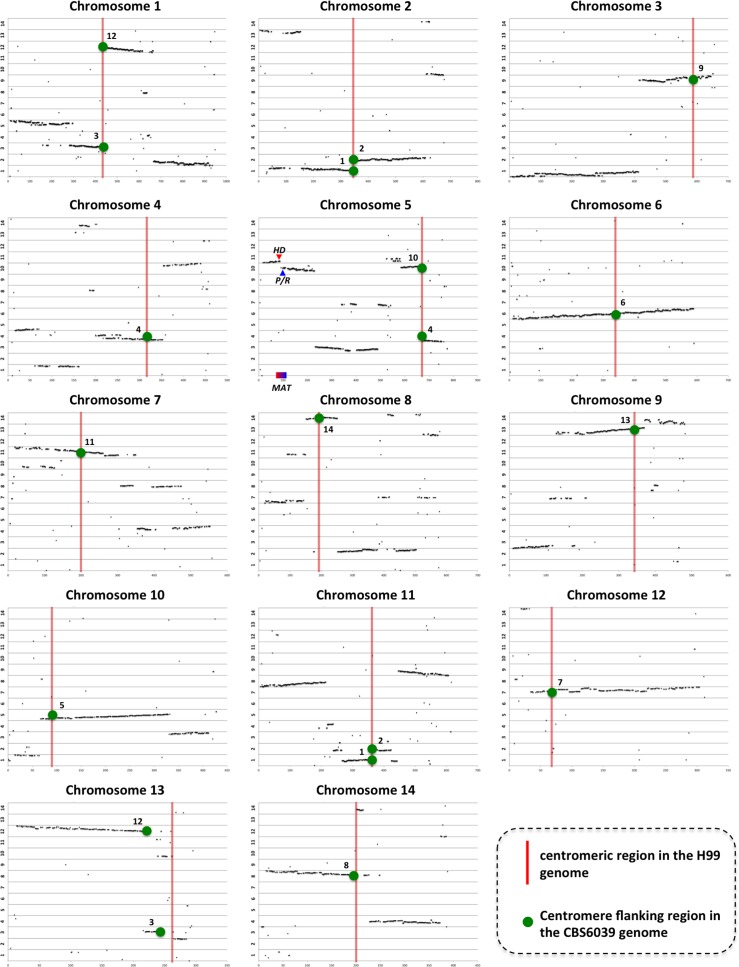
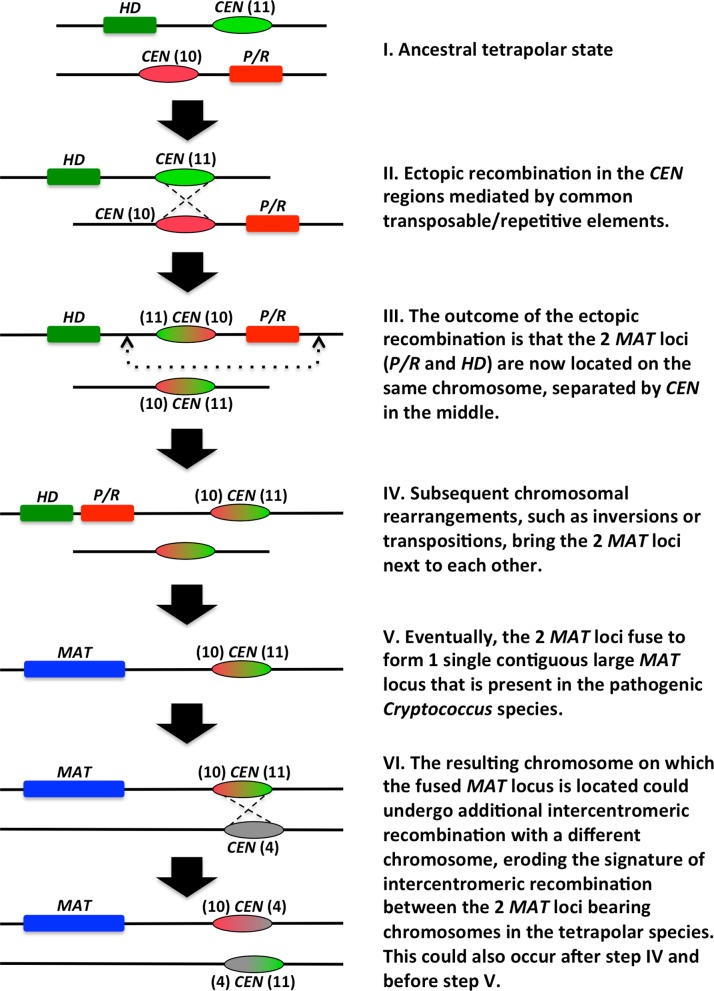
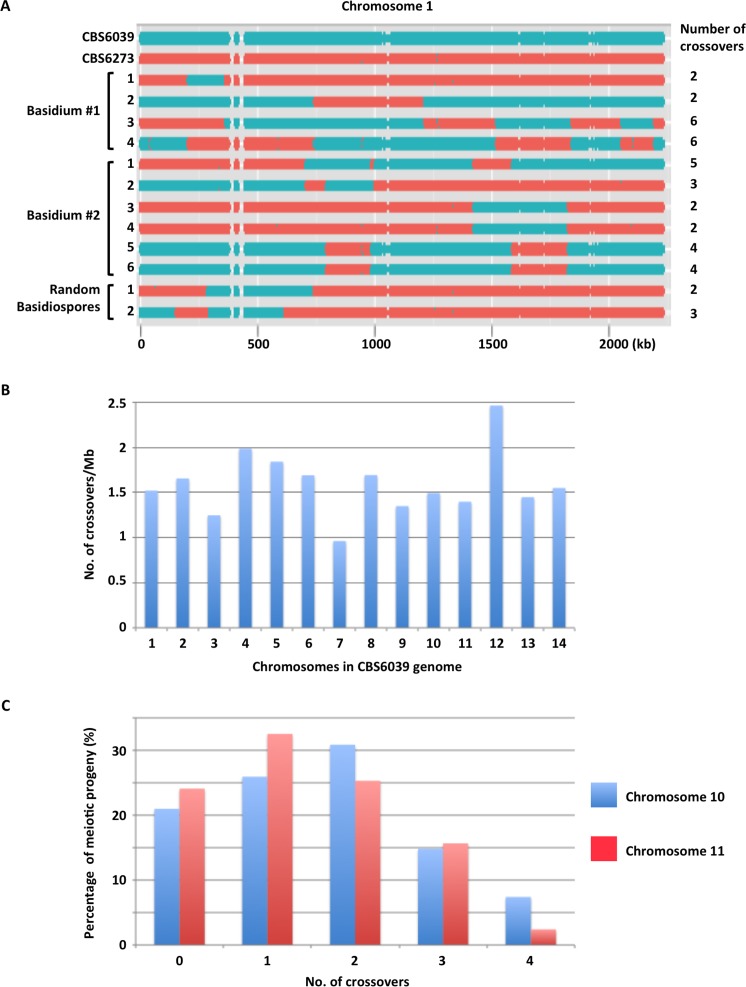
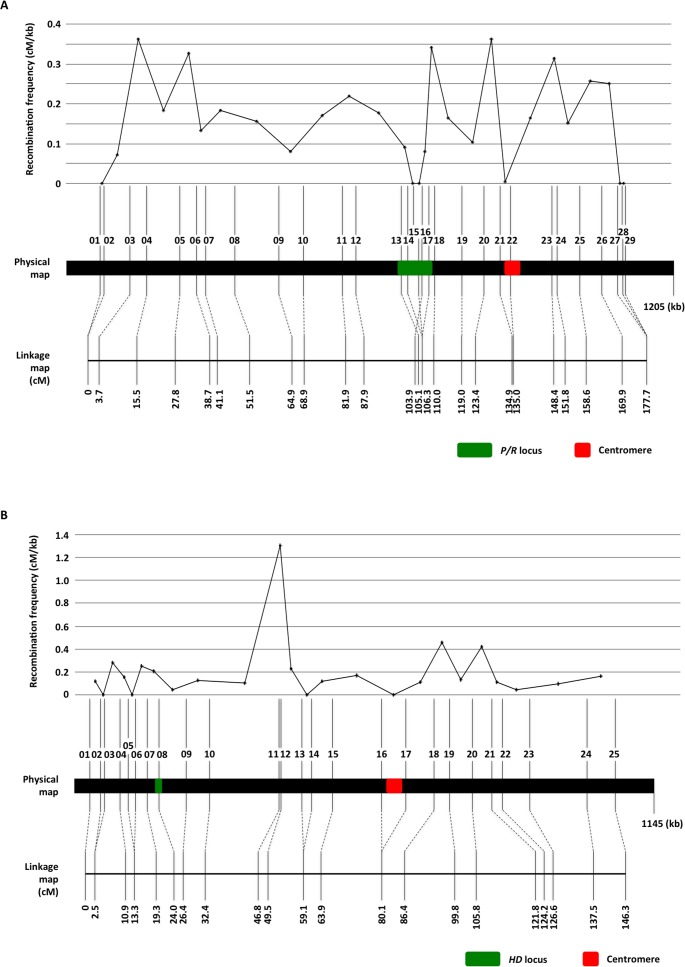
Species within the human pathogenic Cryptococcus species complex are major threats to public health, causing approximately 1 million annual infections globally. Cryptococcus amylolentus is the most closely known related species of the pathogenic Cryptococcus species complex, and it is non-pathogenic. Additionally, while pathogenic Cryptococcus species have bipolar mating systems with a single large mating type (MAT) locus that represents a derived state in Basidiomycetes, C. amylolentus has a tetrapolar mating system with 2 MAT loci (P/R and HD) located on different chromosomes. Thus, studying C. amylolentus will shed light on the transition from tetrapolar to bipolar mating systems in the pathogenic Cryptococcus species, as well as its possible link with the origin and evolution of pathogenesis. In this study, we sequenced, assembled, and annotated the genomes of 2 C. amylolentus isolates, CBS6039 and CBS6273, which are sexual and interfertile. Genome comparison between the 2 C. amylolentus isolates identified the boundaries and the complete gene contents of the P/R and HD MAT loci. Bioinformatic and chromatin immunoprecipitation sequencing (ChIP-seq) analyses revealed that, similar to those of the pathogenic Cryptococcus species, C. amylolentus has regional centromeres (CENs) that are enriched with species-specific transposable and repetitive DNA elements. Additionally, we found that while neither the P/R nor the HD locus is physically closely linked to its centromere in C. amylolentus, and the regions between the MAT loci and their respective centromeres show overall synteny between the 2 genomes, both MAT loci exhibit genetic linkage to their respective centromere during meiosis, suggesting the presence of recombinational suppressors and/or epistatic gene interactions in the MAT-CEN intervening regions. Furthermore, genomic comparisons between C. amylolentus and related pathogenic Cryptococcus species provide evidence that multiple chromosomal rearrangements mediated by intercentromeric recombination have occurred during descent of the 2 lineages from their common ancestor. Taken together, our findings support a model in which the evolution of the bipolar mating system was initiated by an ectopic recombination event mediated by similar repetitive centromeric DNA elements shared between chromosomes. This translocation brought the P/R and HD loci onto the same chromosome, and further chromosomal rearrangements then resulted in the 2 MAT loci becoming physically linked and eventually fusing to form the single contiguous MAT locus that is now extant in the pathogenic Cryptococcus species.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Raper JR. Genetics of sexuality in higher fungi New York: Ronald Press Co.; 1966.
-
- Heitman J, Sun S, James TY. Evolution of fungal sexual reproduction. Mycologia. 2013;105(1):1–27. doi: 10.3852/12-253 - DOI - PubMed
-
- Ni M, Feretzaki M, Sun S, Wang X, Heitman J. Sex in fungi. Annu Rev Genet. 2011;45(1):405–30. doi: 10.1146/annurev-genet-110410-132536 - DOI - PMC - PubMed
-
- Rabe F, Bosch J, Stirnberg A, Guse T, Bauer L, Seitner D, et al. A complete toolset for the study of Ustilago bromivora and Brachypodium sp. as a fungal-temperate grass pathosystem. eLife. 2016;5:e20522 doi: 10.7554/eLife.20522 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
