

The PI3K Pathway in Human Disease
- PMID: 28802037
- PMCID: PMC5726441
- DOI: 10.1016/j.cell.2017.07.029
The PI3K Pathway in Human Disease
Abstract
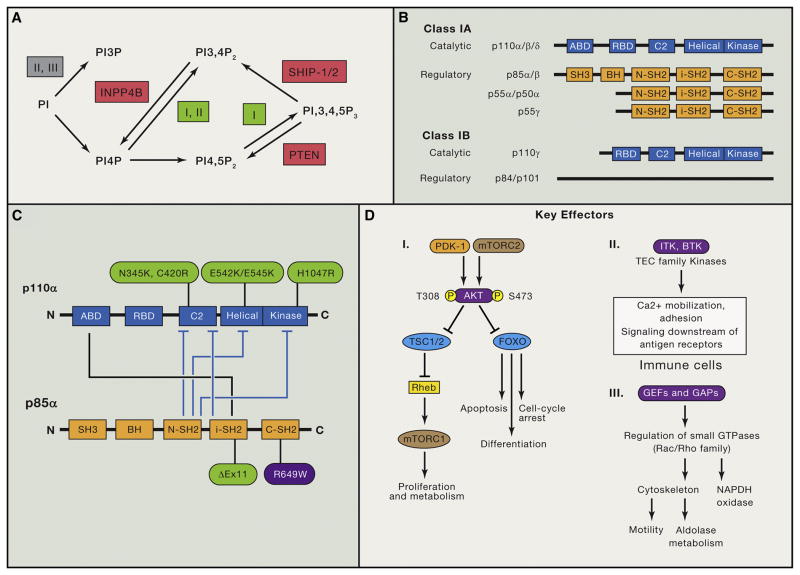
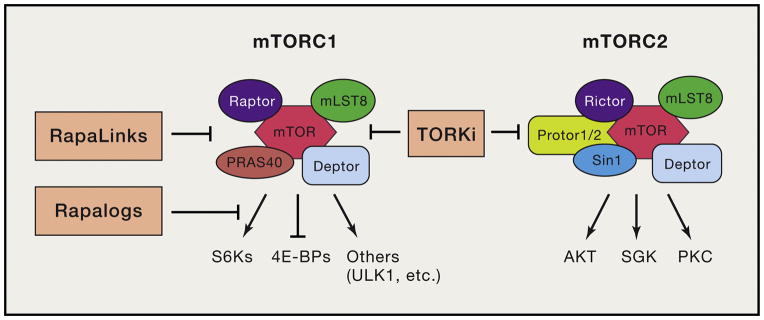
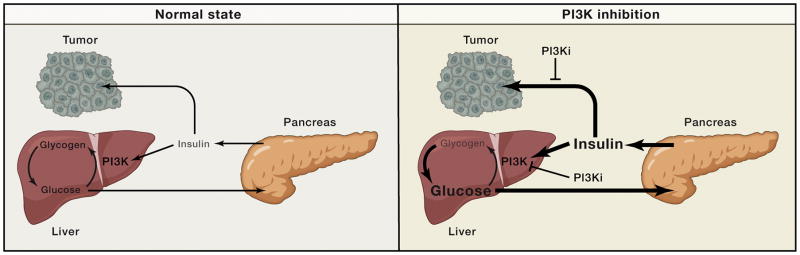
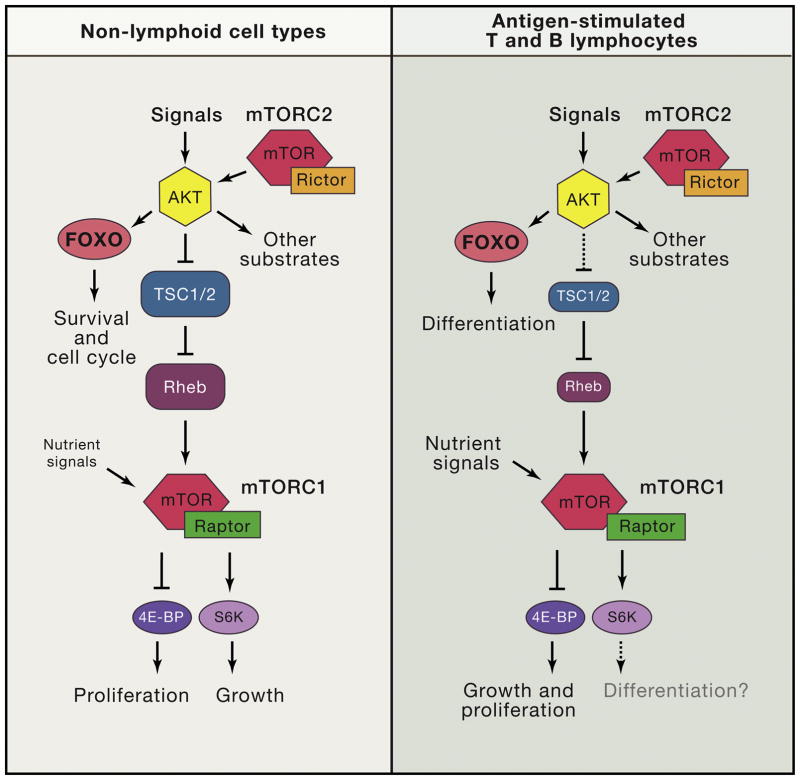
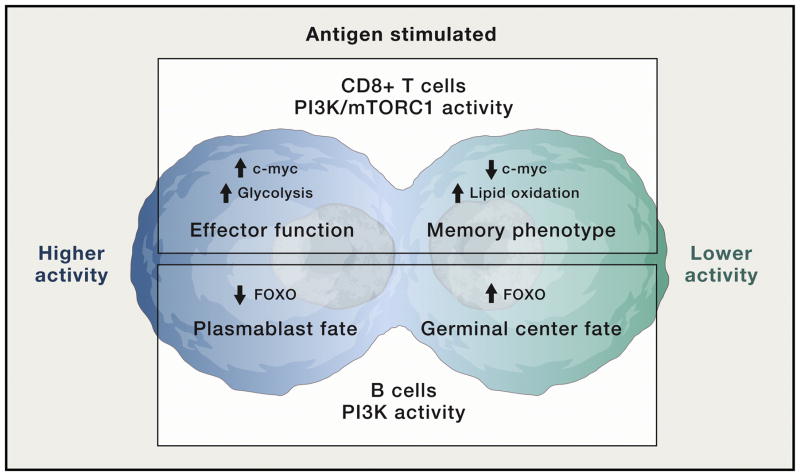
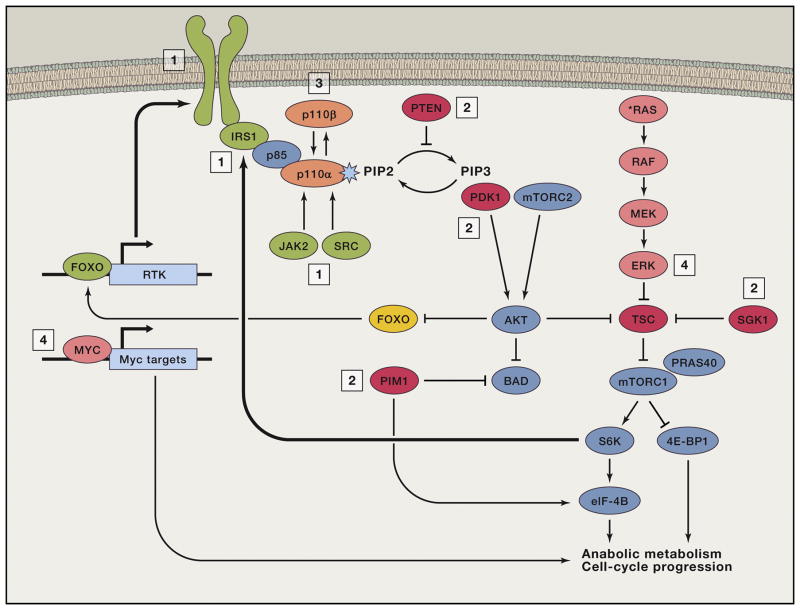
Phosphoinositide 3-kinase (PI3K) activity is stimulated by diverse oncogenes and growth factor receptors, and elevated PI3K signaling is considered a hallmark of cancer. Many PI3K pathway-targeted therapies have been tested in oncology trials, resulting in regulatory approval of one isoform-selective inhibitor (idelalisib) for treatment of certain blood cancers and a variety of other agents at different stages of development. In parallel to PI3K research by cancer biologists, investigations in other fields have uncovered exciting and often unpredicted roles for PI3K catalytic and regulatory subunits in normal cell function and in disease. Many of these functions impinge upon oncology by influencing the efficacy and toxicity of PI3K-targeted therapies. Here we provide a perspective on the roles of class I PI3Ks in the regulation of cellular metabolism and in immune system functions, two topics closely intertwined with cancer biology. We also discuss recent progress developing PI3K-targeted therapies for treatment of cancer and other diseases.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
References
-
- Ahmad S, Abu-Eid R, Shrimali R, Webb M, Verma V, Doroodchi A, Berrong Z, Samara R, Rodriguez PC, Mkrtichyan M, et al. Differential PI3Kδ Signaling in CD4(+) T-cell Subsets Enables Selective Targeting of T Regulatory Cells to Enhance Cancer Immunotherapy. Cancer Res. 2017;77:1892–1904. - PubMed
-
- Aksoy E, Taboubi S, Torres D, Delbauve S, Hachani A, Whitehead MA, Pearce WP, Berenjeno IM, Nock G, Filloux A, et al. The p110δ isoform of the kinase PI(3)K controls the subcellular compartmentalization of TLR4 signaling and protects from endotoxic shock. Nat Immunol. 2012;13:1045–1054. - PMC - PubMed
-
- Ali K, Bilancio A, Thomas M, Pearce W, Gilfillan AM, Tkaczyk C, Kuehn N, Gray A, Giddings J, Peskett E, et al. Essential role for the p110delta phosphoinositide 3-kinase in the allergic response. Nature. 2004;431:1007–1011. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
