

Myc Regulates Chromatin Decompaction and Nuclear Architecture during B Cell Activation
- PMID: 28803781
- PMCID: PMC5854204
- DOI: 10.1016/j.molcel.2017.07.013
Myc Regulates Chromatin Decompaction and Nuclear Architecture during B Cell Activation
Abstract
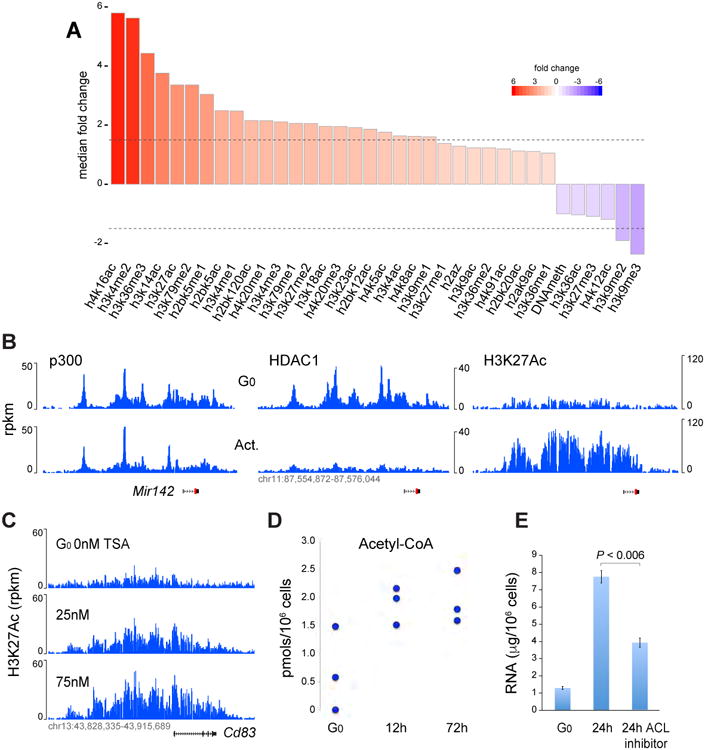
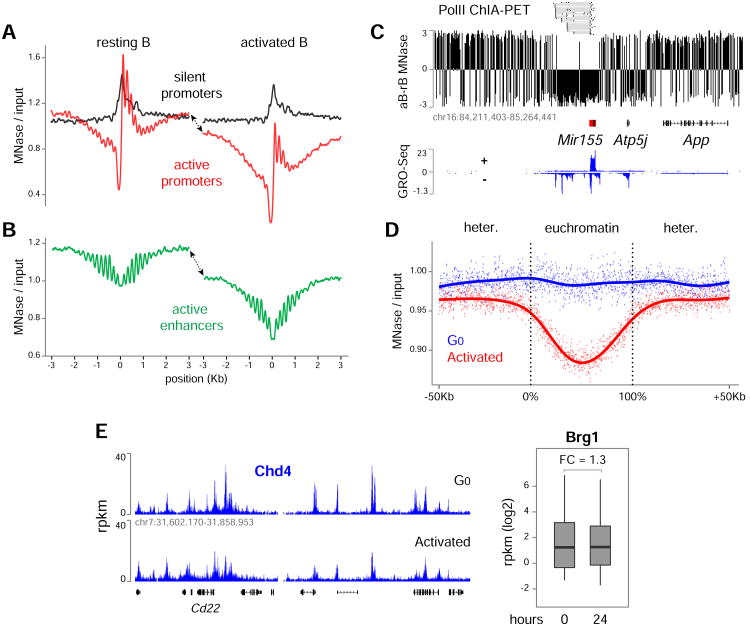
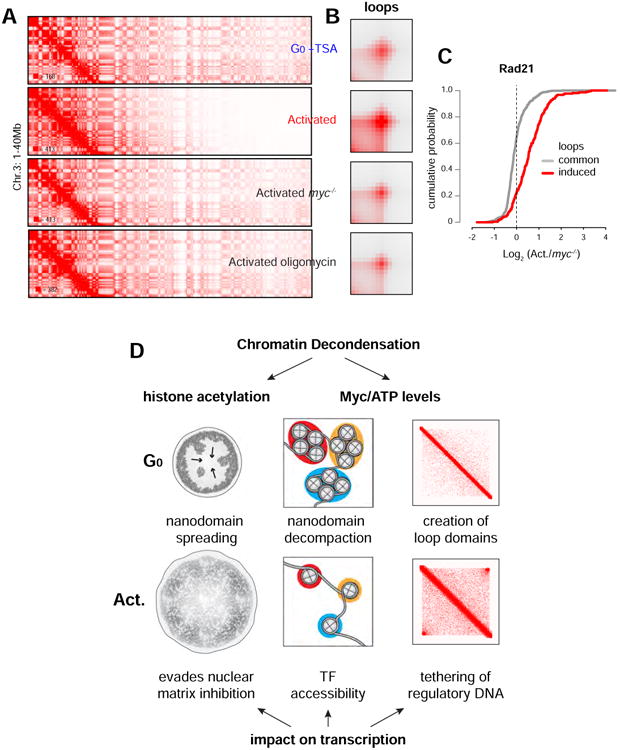
50 years ago, Vincent Allfrey and colleagues discovered that lymphocyte activation triggers massive acetylation of chromatin. However, the molecular mechanisms driving epigenetic accessibility are still unknown. We here show that stimulated lymphocytes decondense chromatin by three differentially regulated steps. First, chromatin is repositioned away from the nuclear periphery in response to global acetylation. Second, histone nanodomain clusters decompact into mononucleosome fibers through a mechanism that requires Myc and continual energy input. Single-molecule imaging shows that this step lowers transcription factor residence time and non-specific collisions during sampling for DNA targets. Third, chromatin interactions shift from long range to predominantly short range, and CTCF-mediated loops and contact domains double in numbers. This architectural change facilitates cognate promoter-enhancer contacts and also requires Myc and continual ATP production. Our results thus define the nature and transcriptional impact of chromatin decondensation and reveal an unexpected role for Myc in the establishment of nuclear topology in mammalian cells.
Keywords: B cells; CTCF; Histone acetylation; chromatin remodeling; cohesin; immune response; myc; nanoscopy; nuclear architecture; transcriptome amplification.
Published by Elsevier Inc.
Conflict of interest statement
Figures




References
-
- Blair D, Dufort FJ, Chiles TC. Protein kinase Cbeta is critical for the metabolic switch to glycolysis following B-cell antigen receptor engagement. Biochem J. 2012;448:165–169. - PubMed
-
- Boggs PT, Donaldson JR, Byrd RH, Schnabel RB. Algorithm 676: software for weighted orthogonal distance regression. ACM Transactions on Mathematical Software (TOMS) 1989;15
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
