

Lengths of the C-Terminus and Interconnecting Loops Impact Stability of Spider-Derived Gating Modifier Toxins
- PMID: 28805686
- PMCID: PMC5577582
- DOI: 10.3390/toxins9080248
Lengths of the C-Terminus and Interconnecting Loops Impact Stability of Spider-Derived Gating Modifier Toxins
Abstract
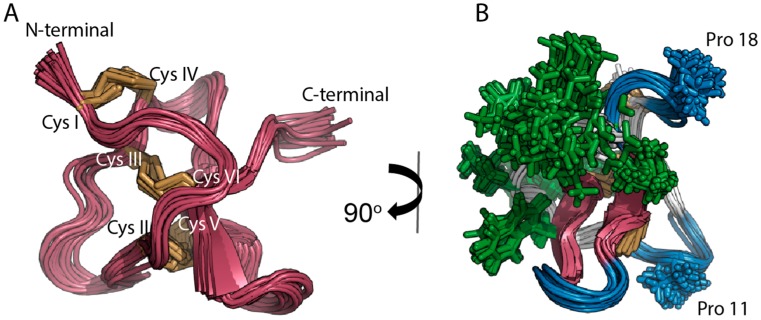
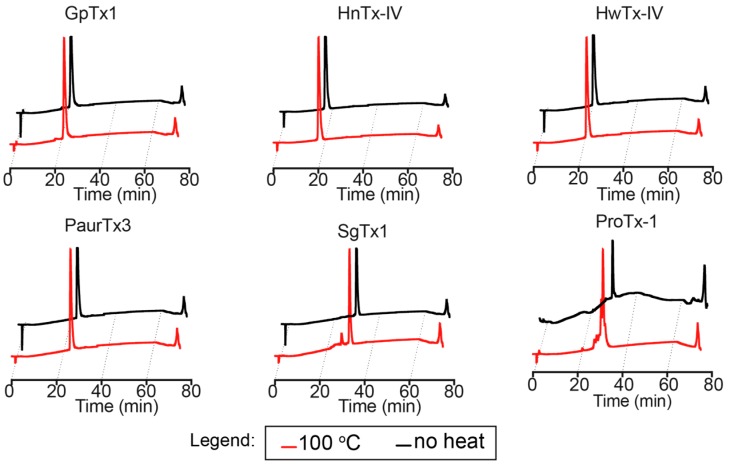
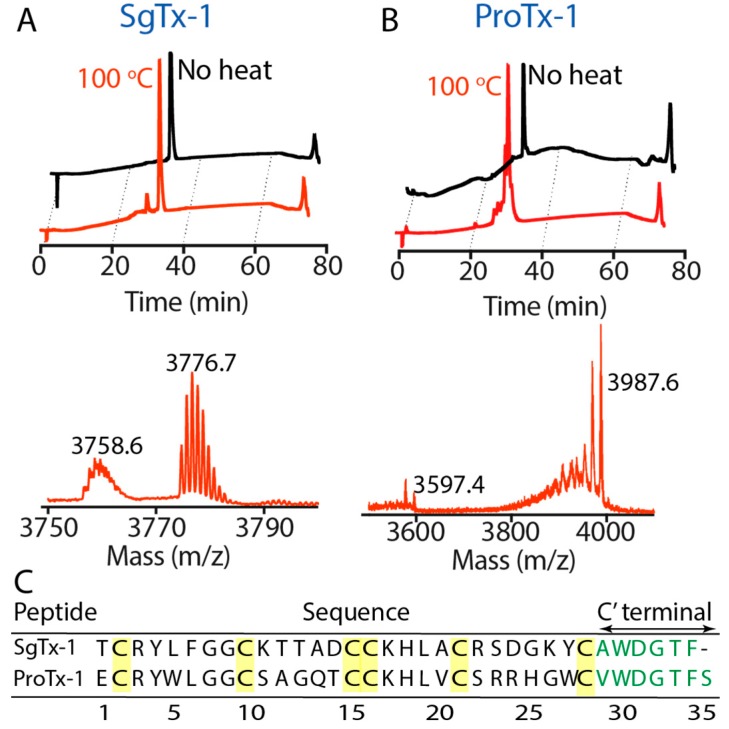
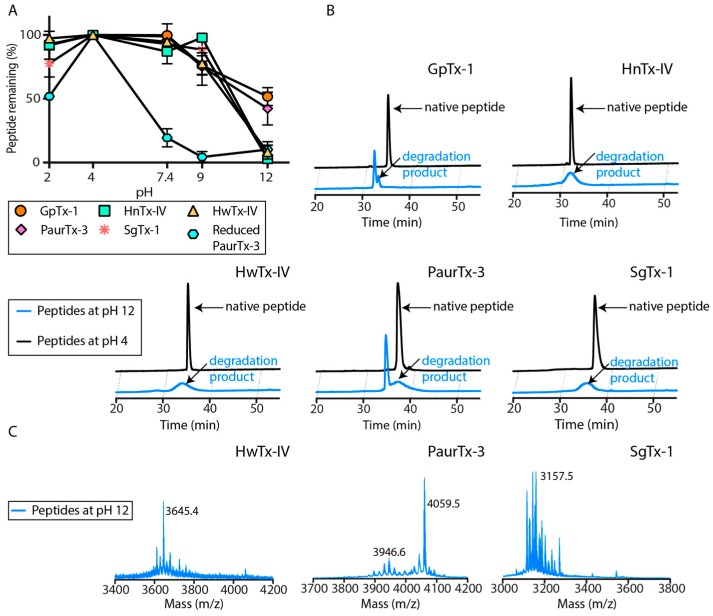
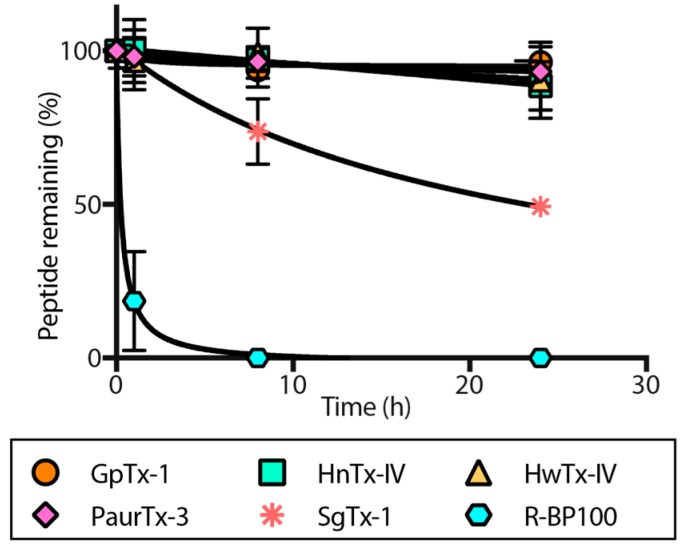
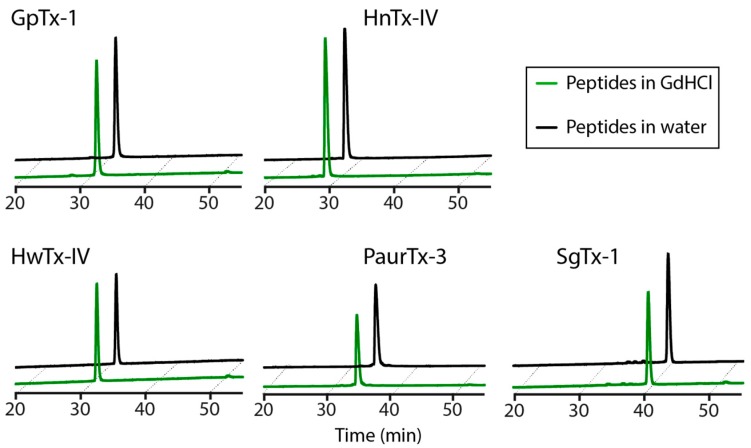
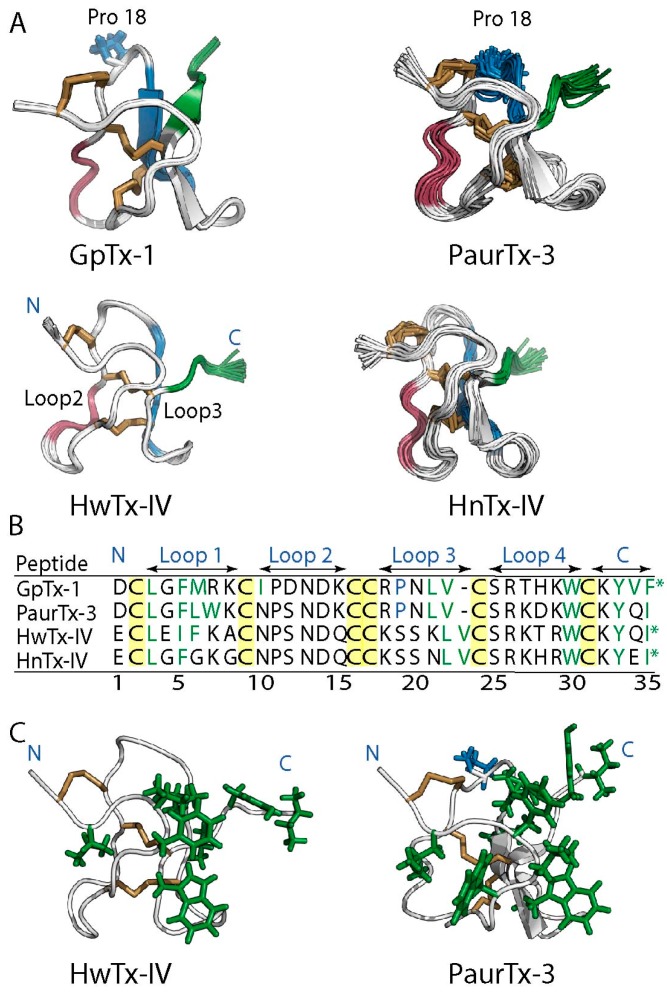
Spider gating modifier toxins (GMTs) are potent modulators of voltage-gated ion channels and have thus attracted attention as drug leads for several pathophysiological conditions. GMTs contain three disulfide bonds organized in an inhibitory cystine knot, which putatively confers them with high stability; however, thus far, there has not been a focused study to establish the stability of GMTs in physiological conditions. We examined the resistance of five GMTs including GpTx-1, HnTx-IV, HwTx-IV, PaurTx-3 and SgTx-1, to pH, thermal and proteolytic degradation. The peptides were stable under physiological conditions, except SgTx-1, which was susceptible to proteolysis, probably due to a longer C-terminus compared to the other peptides. In non-physiological conditions, the five peptides withstood chaotropic degradation, and all but SgTx-1 remained intact after prolonged exposure to high temperature; however, the peptides were degraded in strongly alkaline solutions. GpTx-1 and PaurTx-3 were more resistant to basic hydrolysis than HnTx-IV, HwTx-IV and SgTx-1, probably because a shorter interconnecting loop 3 on GpTx-1 and PaurTx-3 may stabilize interactions between the C-terminus and the hydrophobic patch. Here, we establish that most GMTs are exceptionally stable, and propose that, in the design of GMT-based therapeutics, stability can be enhanced by optimizing the C-terminus in terms of length, and increased interactions with the hydrophobic patch.
Keywords: NaV1.7; nuclear magnetic resonance; pain; rational drug design; serum stability; spider venom.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures


References
-
- Smith J.J., Lau C.H.Y., Herzig V., Ikonomopoulou M.P., Rash L.D., King G.F. Therapeutic applications of spider-venom peptides. In: King G.F., editor. Venoms to Drugs: Venom as a Source for the Development of Human Therapeutics. Royal Society of Chemistry; London, UK: 2015. pp. 221–244.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
