

T-bet-dependent NKp46+ innate lymphoid cells regulate the onset of TH17-induced neuroinflammation
- PMID: 28805812
- PMCID: PMC5605431
- DOI: 10.1038/ni.3816
T-bet-dependent NKp46+ innate lymphoid cells regulate the onset of TH17-induced neuroinflammation
Erratum in
-
Author Correction: T-bet-dependent NKp46+ innate lymphoid cells regulate the onset of TH17-induced neuroinflammation.Nat Immunol. 2018 Aug;19(8):898. doi: 10.1038/s41590-018-0139-6. Nat Immunol. 2018. PMID: 29959442 Free PMC article.
Abstract
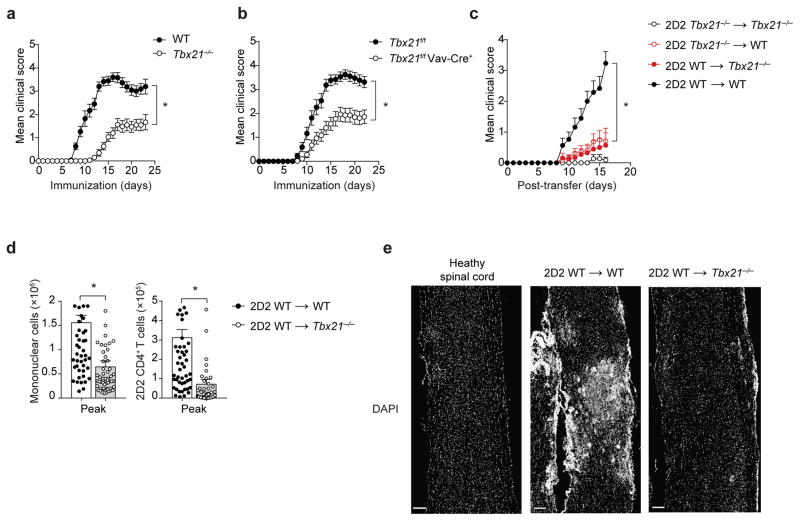
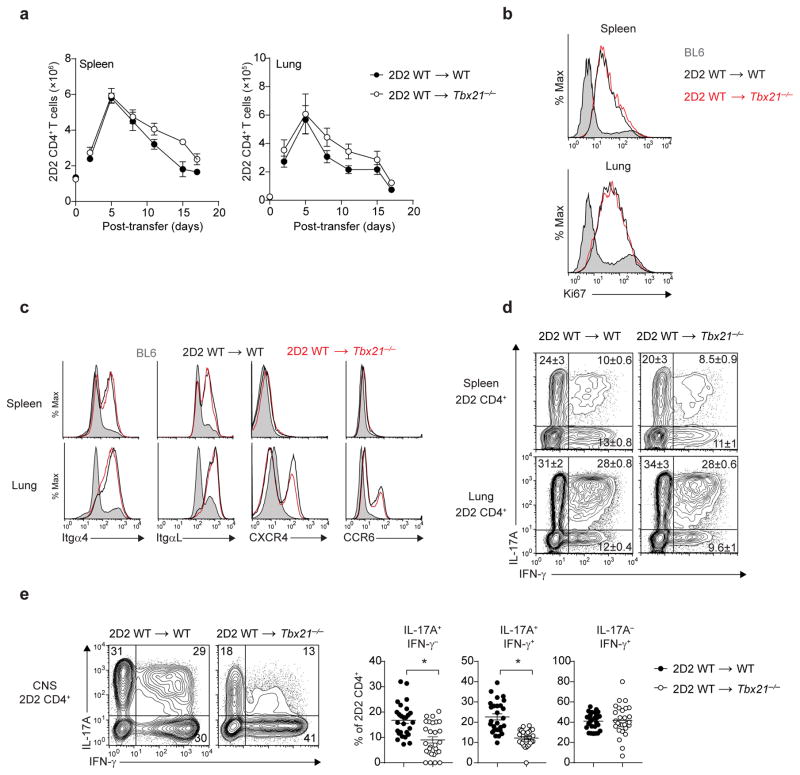
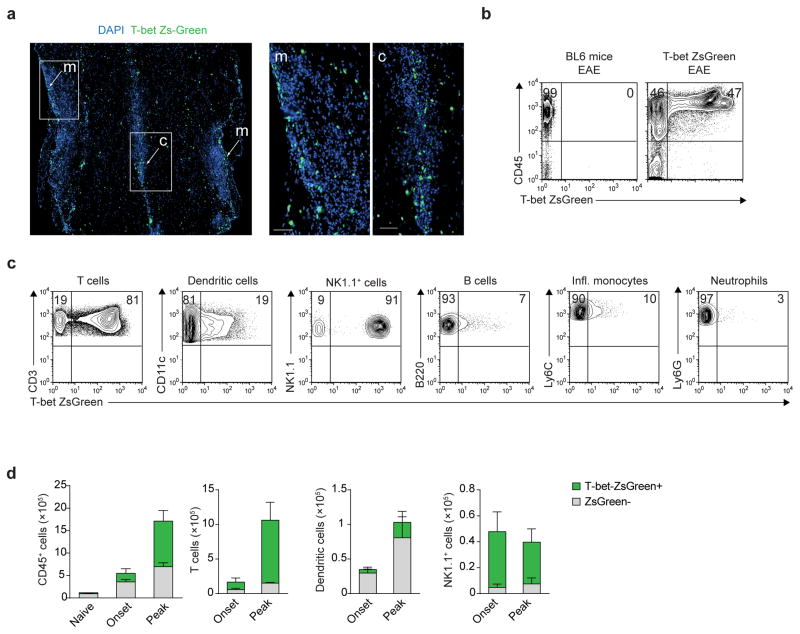
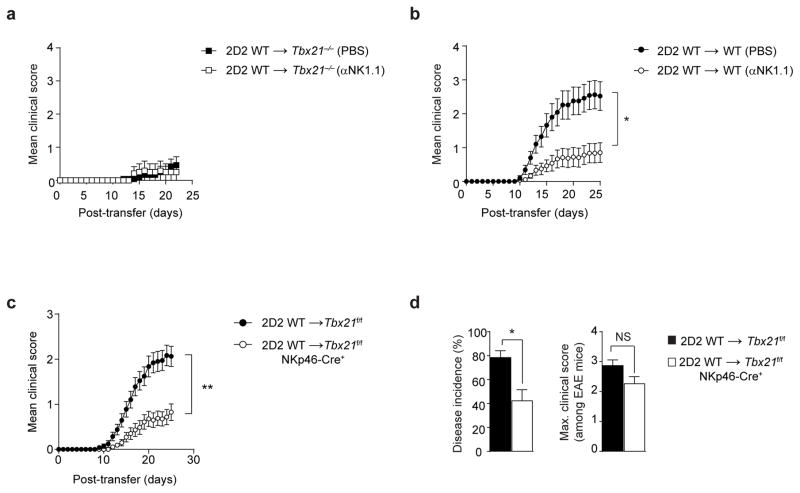
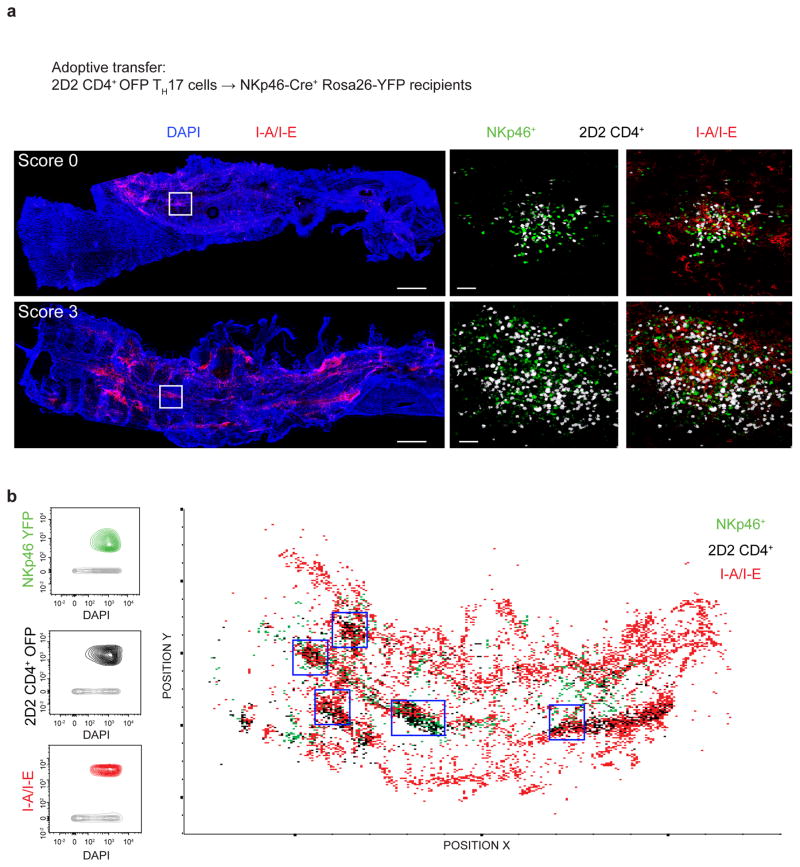
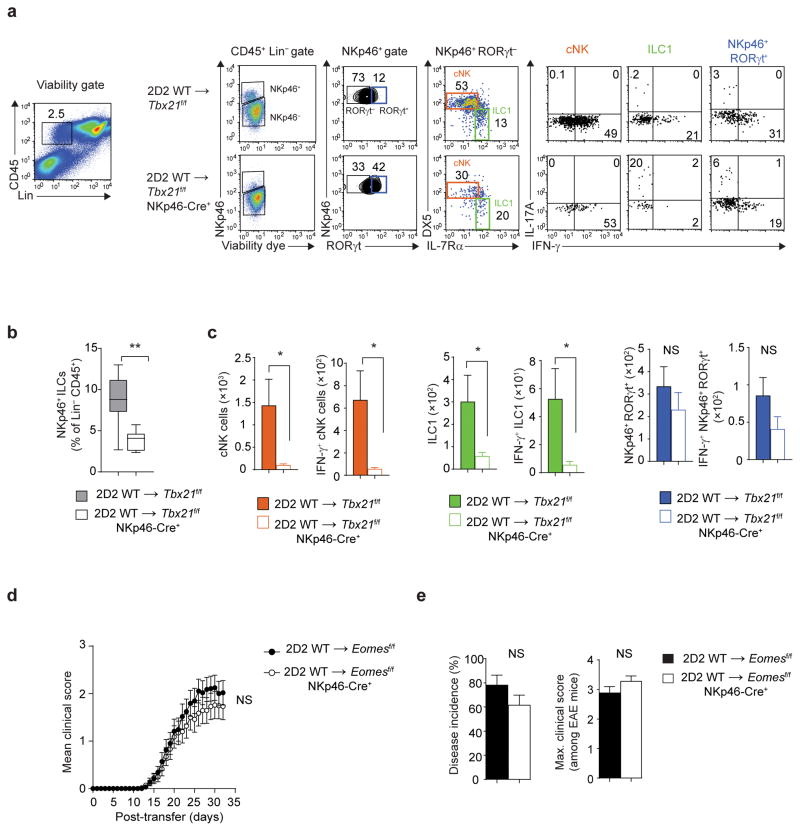
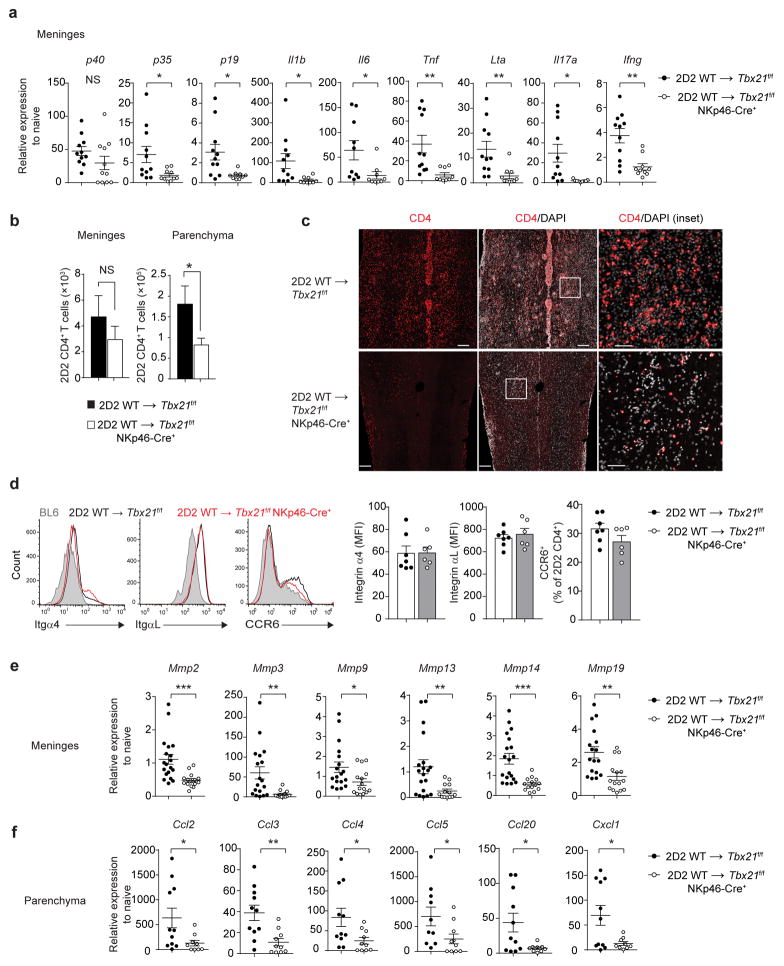
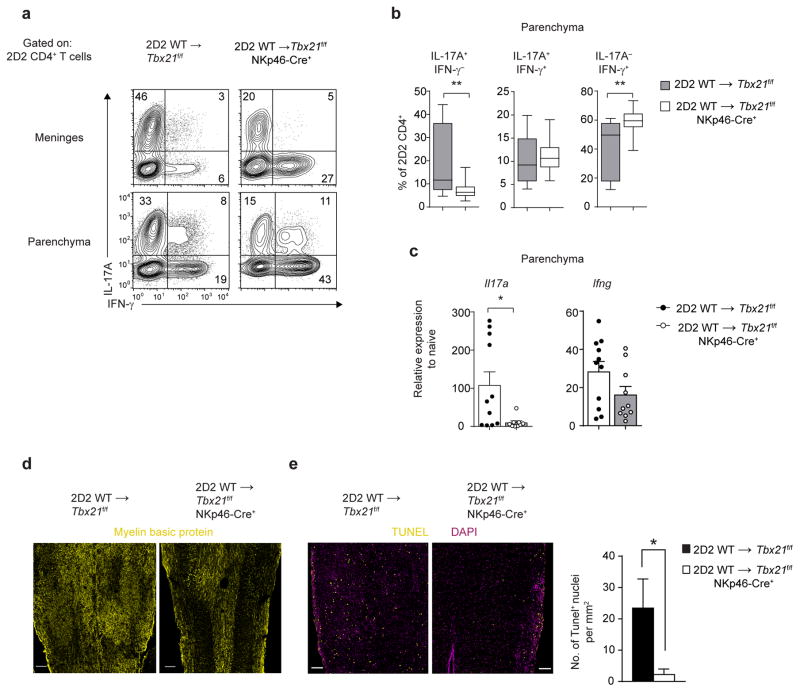
The transcription factor T-bet has been associated with increased susceptibility to systemic and organ-specific autoimmunity, but the mechanism by which T-bet expression promotes neuroinflammation remains unknown. In this study, we demonstrate a cardinal role of T-bet-dependent NKp46+ innate lymphoid cells (ILCs) in the initiation of CD4+ TH17-mediated neuroinflammation. Loss of T-bet specifically in NKp46+ ILCs profoundly impaired the ability of myelin-reactive TH17 cells to invade central nervous system (CNS) tissue and protected the mice from autoimmunity. T-bet-dependent NKp46+ ILCs localized in the meninges and acted as chief coordinators of meningeal inflammation by inducing the expression of proinflammatory cytokines, chemokines and matrix metalloproteinases, which together facilitated T cell entry into CNS parenchyma. Our findings uncover a detrimental role of T-bet-dependent NKp46+ ILCs in the development of CNS autoimmune disease.
Conflict of interest statement
COMPETING FINANCIAL INTERESTS
The authors declare no competing financial interests.
Figures
Comment in
-
(T)Betting on innate lymphoid cells in CNS inflammatory disease.Nat Immunol. 2017 Sep 19;18(10):1063-1064. doi: 10.1038/ni.3839. Nat Immunol. 2017. PMID: 28926546 No abstract available.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
