

A systems approach reveals distinct metabolic strategies among the NCI-60 cancer cell lines
- PMID: 28806730
- PMCID: PMC5570491
- DOI: 10.1371/journal.pcbi.1005698
A systems approach reveals distinct metabolic strategies among the NCI-60 cancer cell lines
Abstract
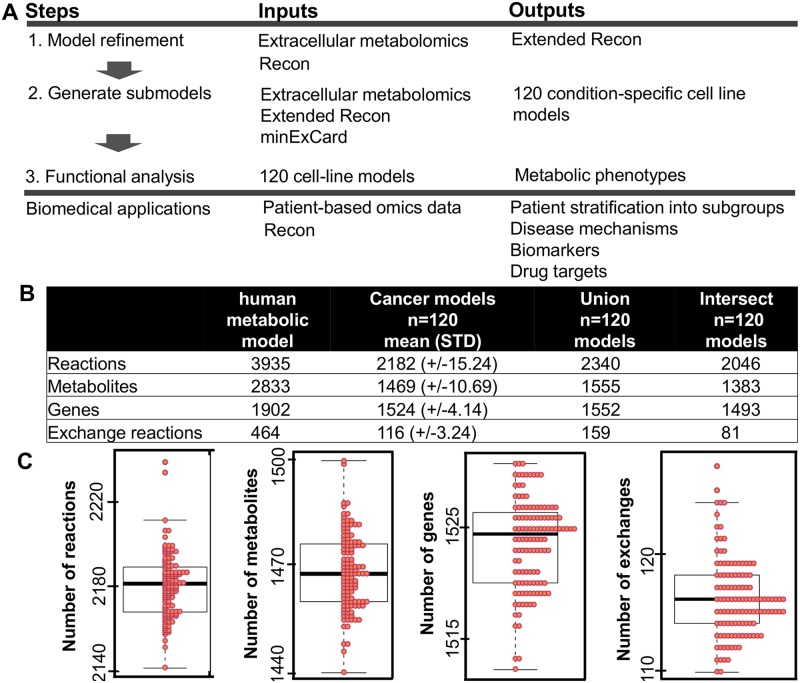
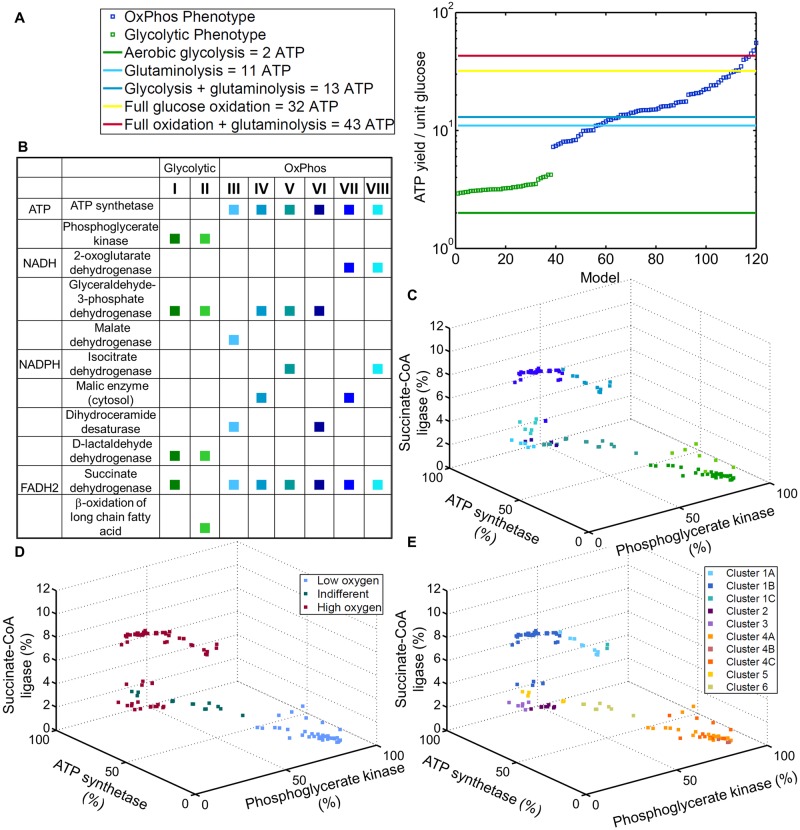
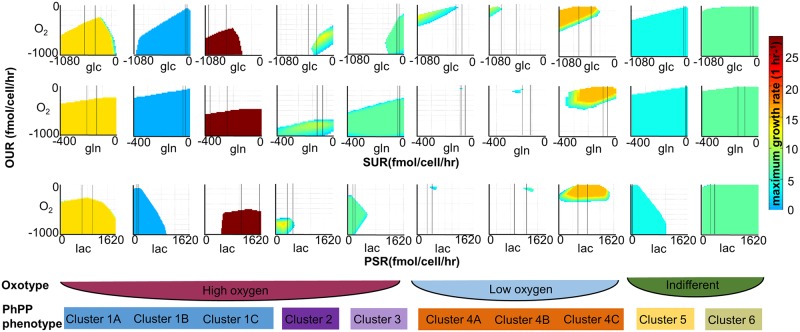
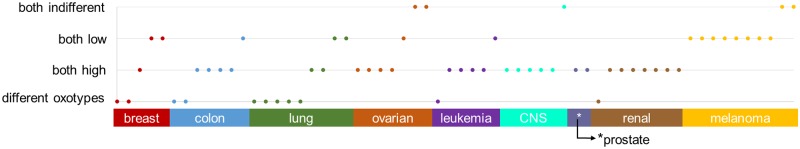
The metabolic phenotype of cancer cells is reflected by the metabolites they consume and by the byproducts they release. Here, we use quantitative, extracellular metabolomic data of the NCI-60 panel and a novel computational method to generate 120 condition-specific cancer cell line metabolic models. These condition-specific cancer models used distinct metabolic strategies to generate energy and cofactors. The analysis of the models' capability to deal with environmental perturbations revealed three oxotypes, differing in the range of allowable oxygen uptake rates. Interestingly, models based on metabolomic profiles of melanoma cells were distinguished from other models through their low oxygen uptake rates, which were associated with a glycolytic phenotype. A subset of the melanoma cell models required reductive carboxylation. The analysis of protein and RNA expression levels from the Human Protein Atlas showed that IDH2, which was an essential gene in the melanoma models, but not IDH1 protein, was detected in normal skin cell types and melanoma. Moreover, the von Hippel-Lindau tumor suppressor (VHL) protein, whose loss is associated with non-hypoxic HIF-stabilization, reductive carboxylation, and promotion of glycolysis, was uniformly absent in melanoma. Thus, the experimental data supported the predicted role of IDH2 and the absence of VHL protein supported the glycolytic and low oxygen phenotype predicted for melanoma. Taken together, our approach of integrating extracellular metabolomic data with metabolic modeling and the combination of different network interrogation methods allowed insights into the metabolism of cells.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
