

PKA activity is essential for relieving the suppression of hyphal growth and appressorium formation by MoSfl1 in Magnaporthe oryzae
- PMID: 28806765
- PMCID: PMC5570492
- DOI: 10.1371/journal.pgen.1006954
PKA activity is essential for relieving the suppression of hyphal growth and appressorium formation by MoSfl1 in Magnaporthe oryzae
Abstract
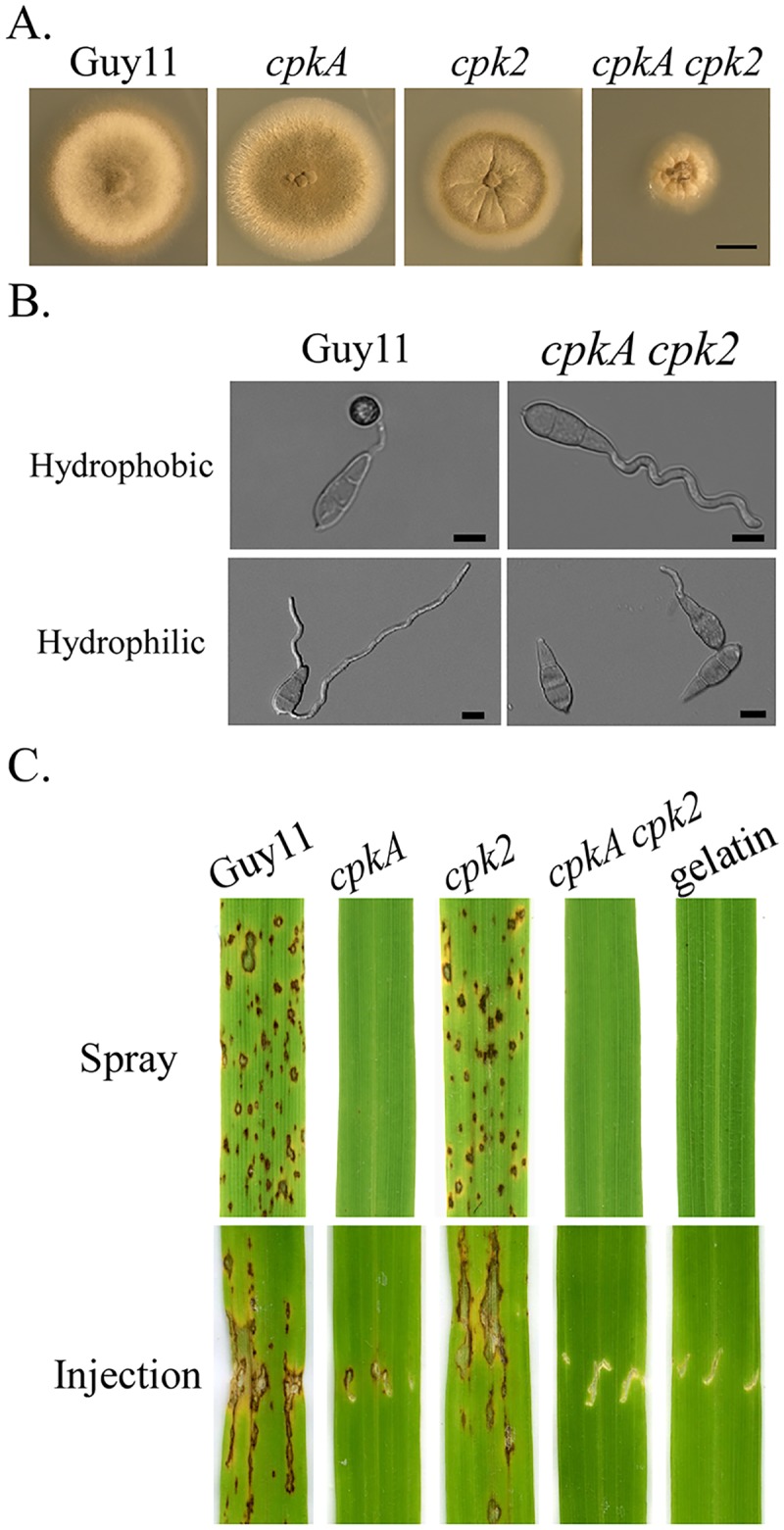
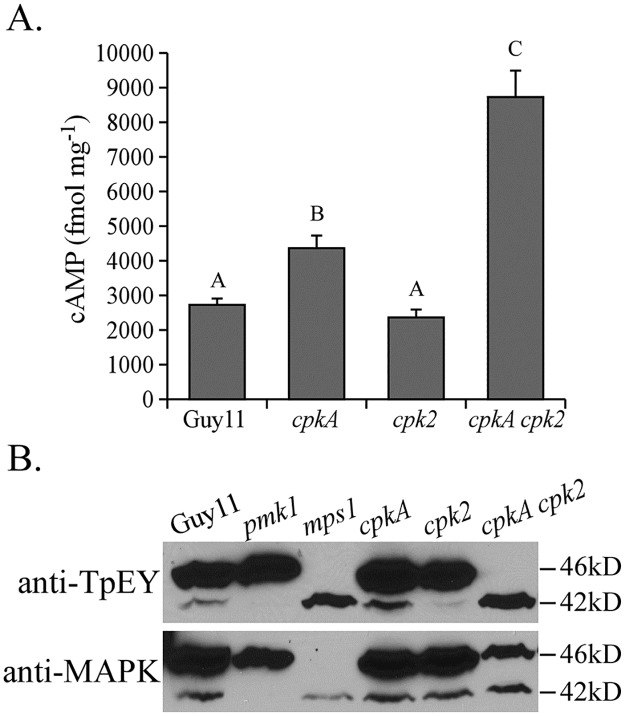
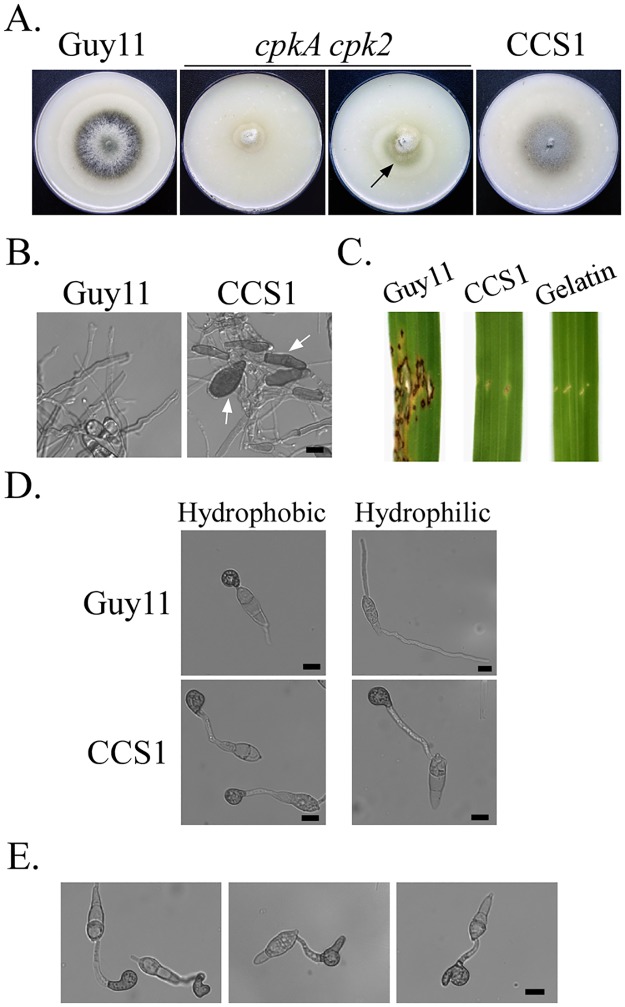
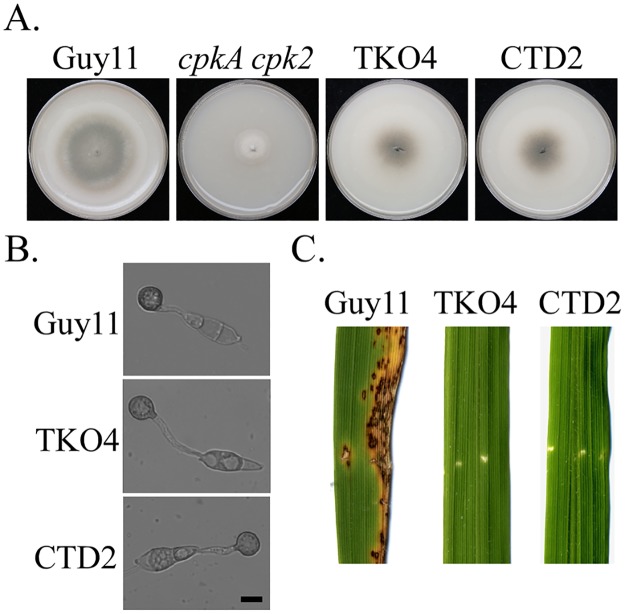
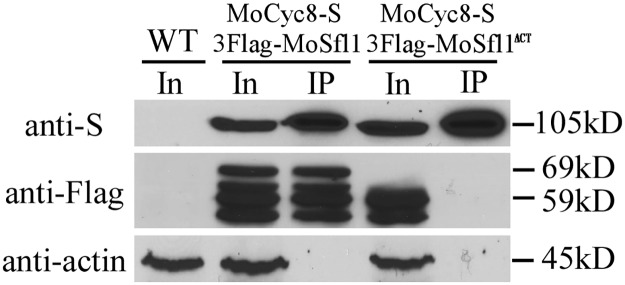
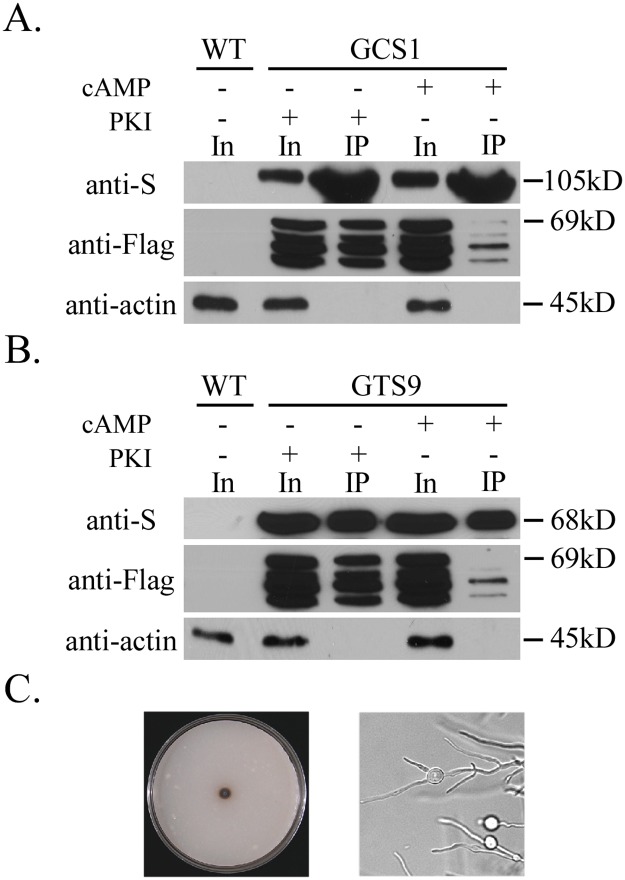
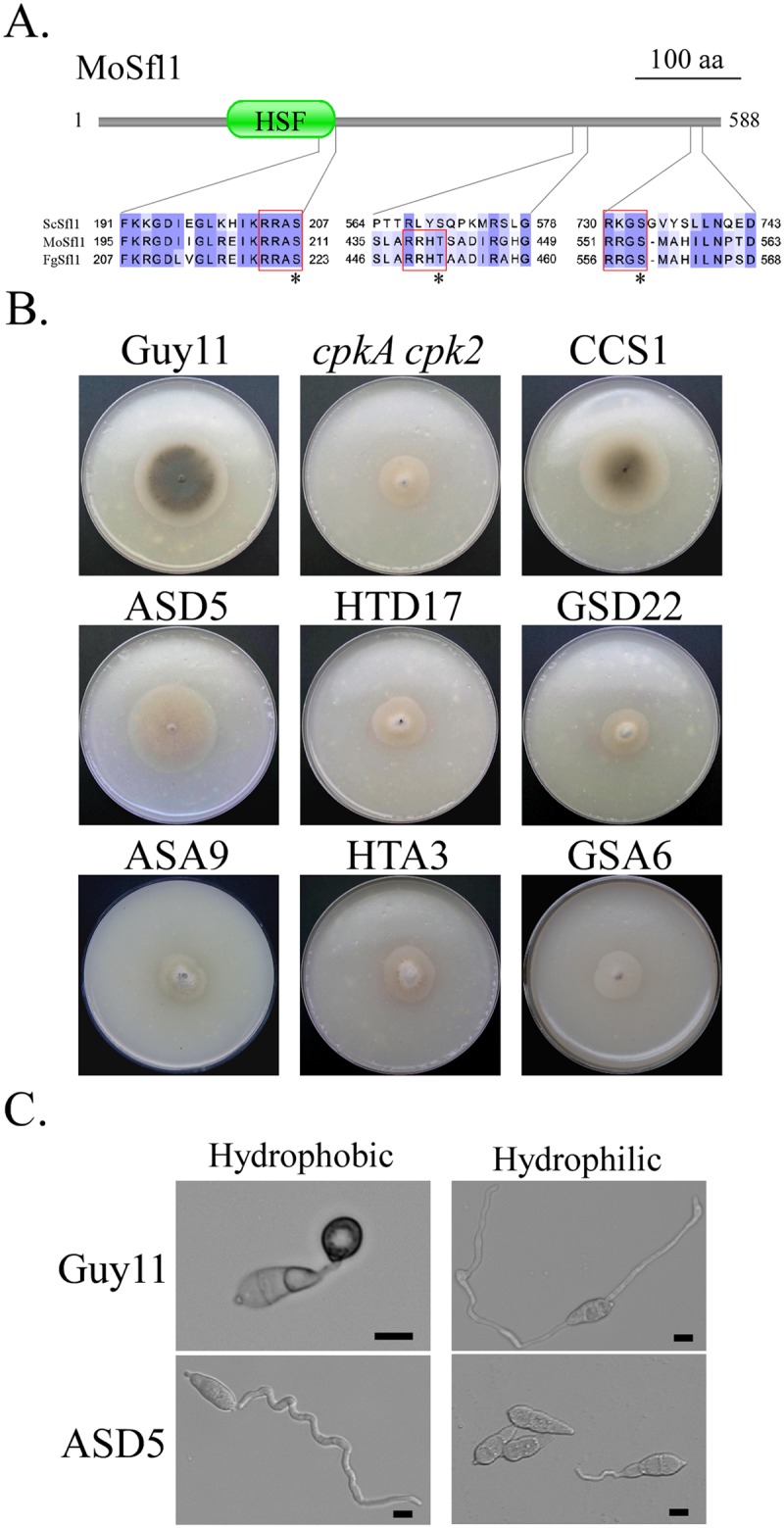
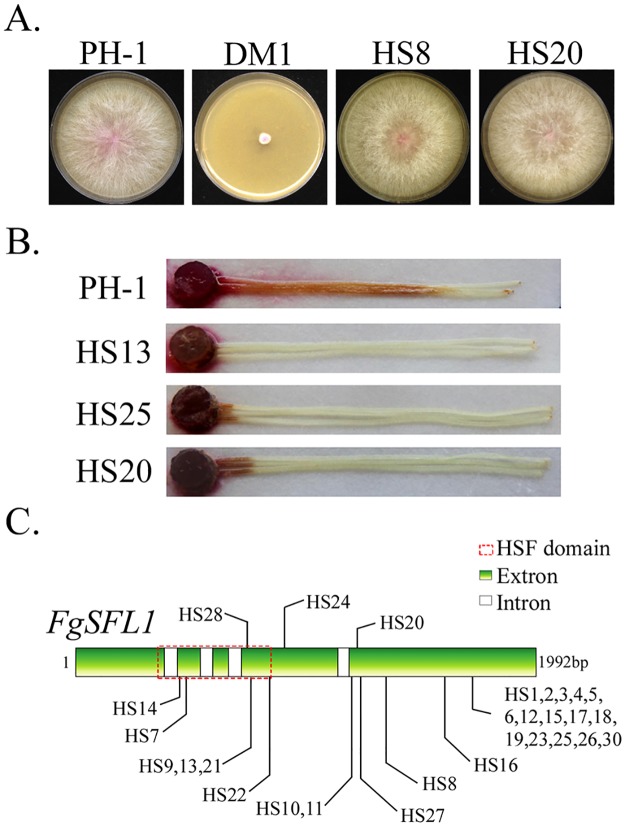
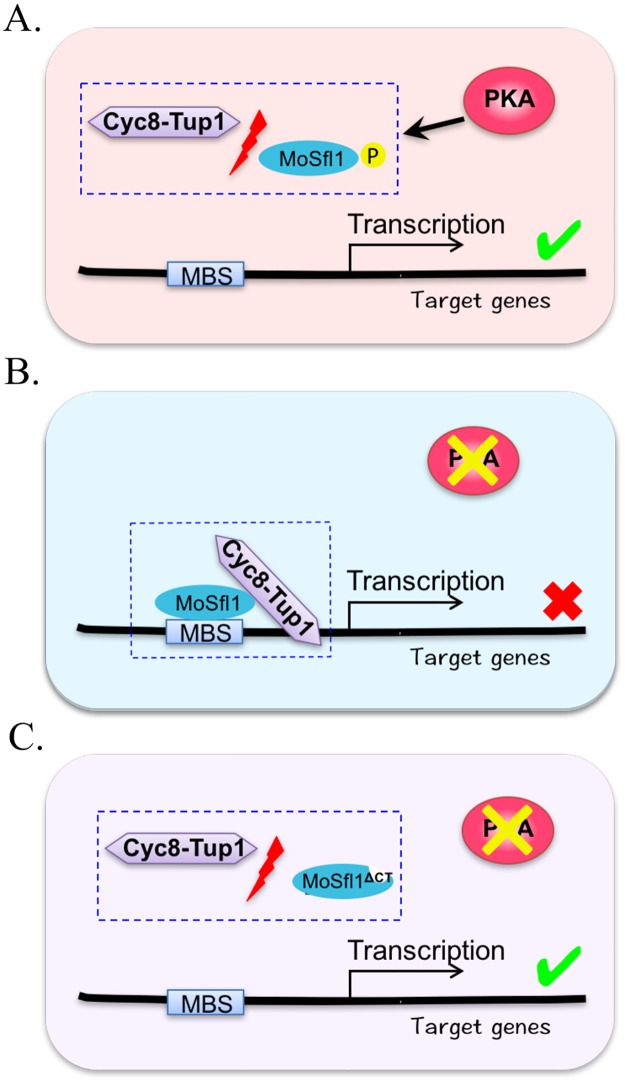
In the rice blast fungus Magnaporthe oryzae, the cAMP-PKA pathway regulates surface recognition, appressorium turgor generation, and invasive growth. However, deletion of CPKA failed to block appressorium formation and responses to exogenous cAMP. In this study, we generated and characterized the cpk2 and cpkA cpk2 mutants and spontaneous suppressors of cpkA cpk2 in M. oryzae. Our results demonstrate that CPKA and CPK2 have specific and overlapping functions, and PKA activity is essential for appressorium formation and plant infection. Unlike the single mutants, the cpkA cpk2 mutant was significantly reduced in growth and rarely produced conidia. It failed to form appressoria although the intracellular cAMP level and phosphorylation of Pmk1 MAP kinase were increased. The double mutant also was defective in plant penetration and Mps1 activation. Interestingly, it often produced fast-growing spontaneous suppressors that formed appressoria but were still non-pathogenic. Two suppressor strains of cpkA cpk2 had deletion and insertion mutations in the MoSFL1 transcription factor gene. Deletion of MoSFL1 or its C-terminal 93-aa (MoSFL1ΔCT) was confirmed to suppress the defects of cpkA cpk2 in hyphal growth but not appressorium formation or pathogenesis. We also isolated 30 spontaneous suppressors of the cpkA cpk2 mutant in Fusarium graminearum and identified mutations in 29 of them in FgSFL1. Affinity purification and co-IP assays showed that this C-terminal region of MoSfl1 was essential for its interaction with the conserved Cyc8-Tup1 transcriptional co-repressor, which was reduced by cAMP treatment. Furthermore, the S211D mutation at the conserved PKA-phosphorylation site in MoSFL1 partially suppressed the defects of cpkA cpk2. Overall, our results indicate that PKA activity is essential for appressorium formation and proper activation of Pmk1 or Mps1 in M. oryzae, and phosphorylation of MoSfl1 by PKA relieves its interaction with the Cyc8-Tup1 co-repressor and suppression of genes important for hyphal growth.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Dean RA, Talbot NJ, Ebbole DJ, Farman ML, Mitchell TK, Orbach MJ, et al. The genome sequence of the rice blast fungus Magnaporthe grisea. Nature. 2005;434(7036):980–6. doi: 10.1038/nature03449 - DOI - PubMed
-
- Wilson RA, Talbot NJ. Under pressure: investigating the biology of plant infection by Magnaporthe oryzae. Nat Rev Microbiol. 2009;7(3):185–95. doi: 10.1038/nrmicro2032 - DOI - PubMed
-
- Xu JR, Zhao X, Dean RA. From genes to genomes: a new paradigm for studying fungal pathogenesis in Magnaporthe oryzae. Adv Genet. 2007;57:175–218. doi: 10.1016/S0065-2660(06)57005-1 - DOI - PubMed
-
- Dagdas YF, Yoshino K, Dagdas G, Ryder LS, Bielska E, Steinberg G, et al. Septin-mediated plant cell invasion by the rice blast fungus, Magnaporthe oryzae. Science. 2012;336(6088):1590–5. doi: 10.1126/science.1222934 - DOI - PubMed
-
- Kankanala P, Czymmek K, Valent B. Roles for rice membrane dynamics and plasmodesmata during biotrophic invasion by the blast fungus. Plant Cell. 2007;19(2):706–24. doi: 10.1105/tpc.106.046300 - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
