

Breath-giving cooperation: critical review of origin of mitochondria hypotheses : Major unanswered questions point to the importance of early ecology
- PMID: 28806979
- PMCID: PMC5557255
- DOI: 10.1186/s13062-017-0190-5
Breath-giving cooperation: critical review of origin of mitochondria hypotheses : Major unanswered questions point to the importance of early ecology
Abstract
The origin of mitochondria is a unique and hard evolutionary problem, embedded within the origin of eukaryotes. The puzzle is challenging due to the egalitarian nature of the transition where lower-level units took over energy metabolism. Contending theories widely disagree on ancestral partners, initial conditions and unfolding of events. There are many open questions but there is no comparative examination of hypotheses. We have specified twelve questions about the observable facts and hidden processes leading to the establishment of the endosymbiont that a valid hypothesis must address. We have objectively compared contending hypotheses under these questions to find the most plausible course of events and to draw insight on missing pieces of the puzzle. Since endosymbiosis borders evolution and ecology, and since a realistic theory has to comply with both domains' constraints, the conclusion is that the most important aspect to clarify is the initial ecological relationship of partners. Metabolic benefits are largely irrelevant at this initial phase, where ecological costs could be more disruptive. There is no single theory capable of answering all questions indicating a severe lack of ecological considerations. A new theory, compliant with recent phylogenomic results, should adhere to these criteria.
Reviewers: This article was reviewed by Michael W. Gray, William F. Martin and Purificación López-García.
Keywords: Ecology; Endosymbiosis; Eukaryogenesis; Evolution; Major transition; Metabolism; Mitochondria; Parasitism.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
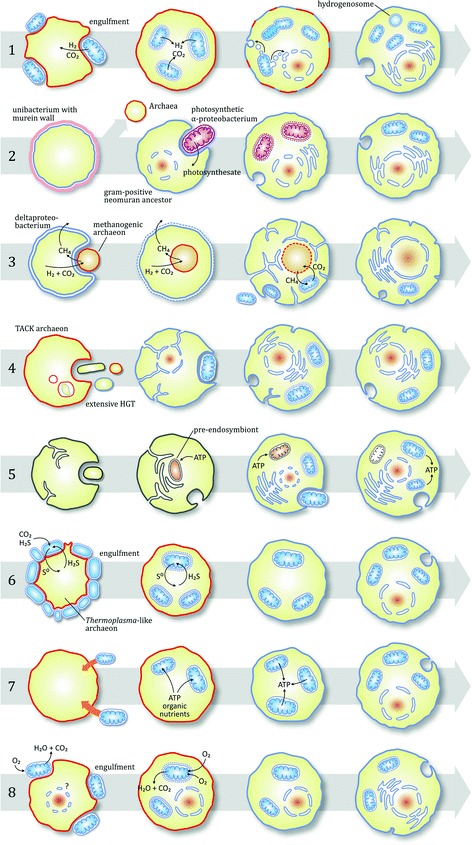
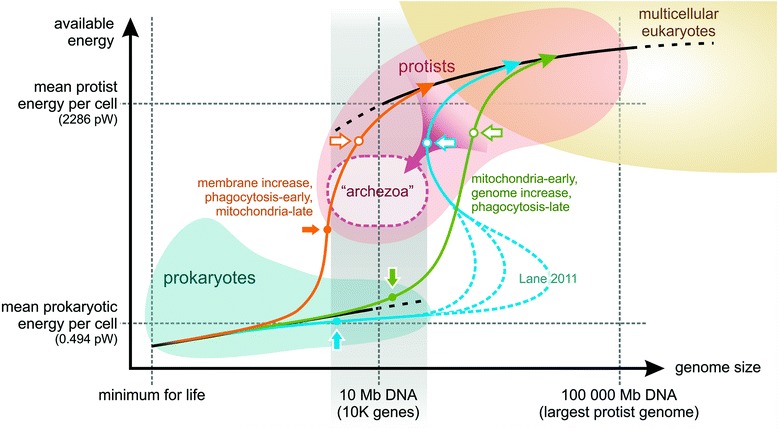
References
-
- Maynard Smith J, Szathmáry E. The major transitions in evolution. Oxford: Freeman & Co.; 1995. p. 360.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
