

Cytoplasmic poly (A)-binding protein critically regulates epidermal maintenance and turnover in the planarian Schmidtea mediterranea
- PMID: 28807897
- PMCID: PMC5611960
- DOI: 10.1242/dev.152942
Cytoplasmic poly (A)-binding protein critically regulates epidermal maintenance and turnover in the planarian Schmidtea mediterranea
Abstract
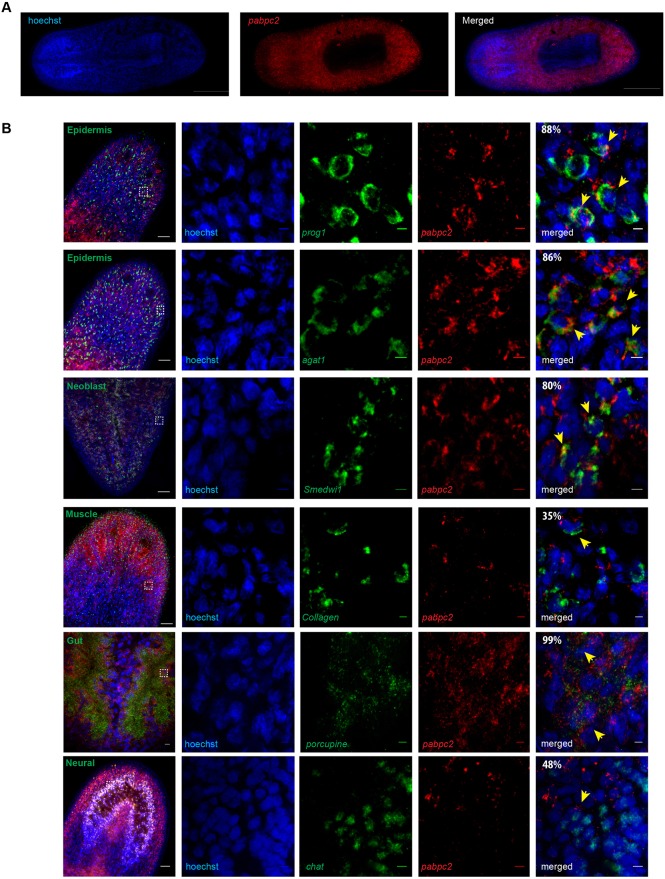
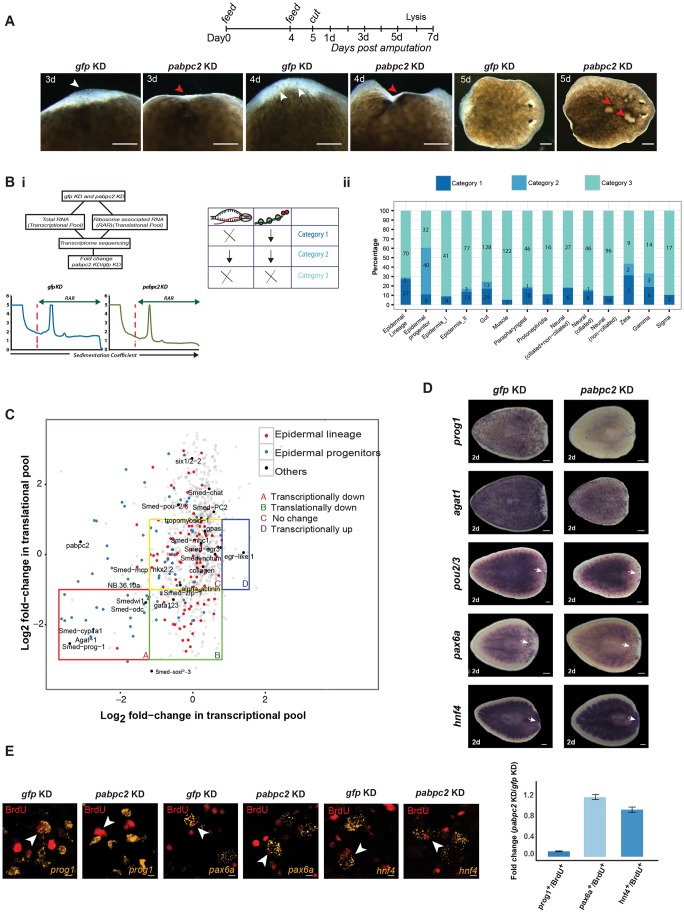
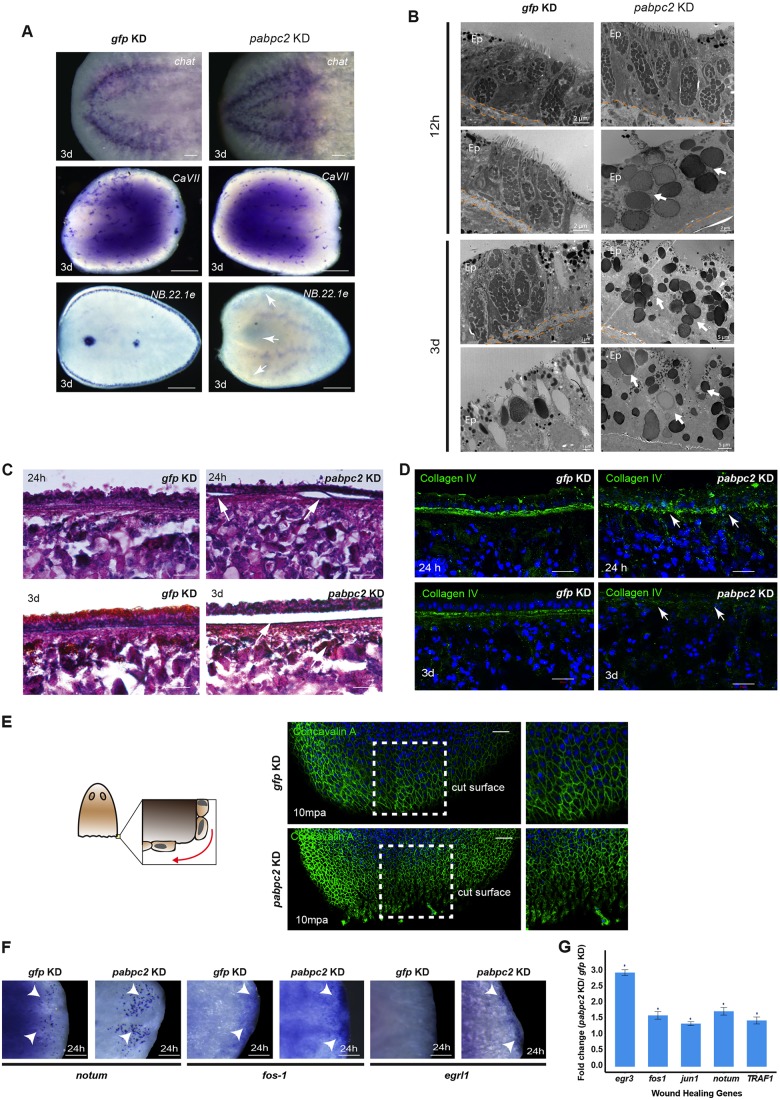
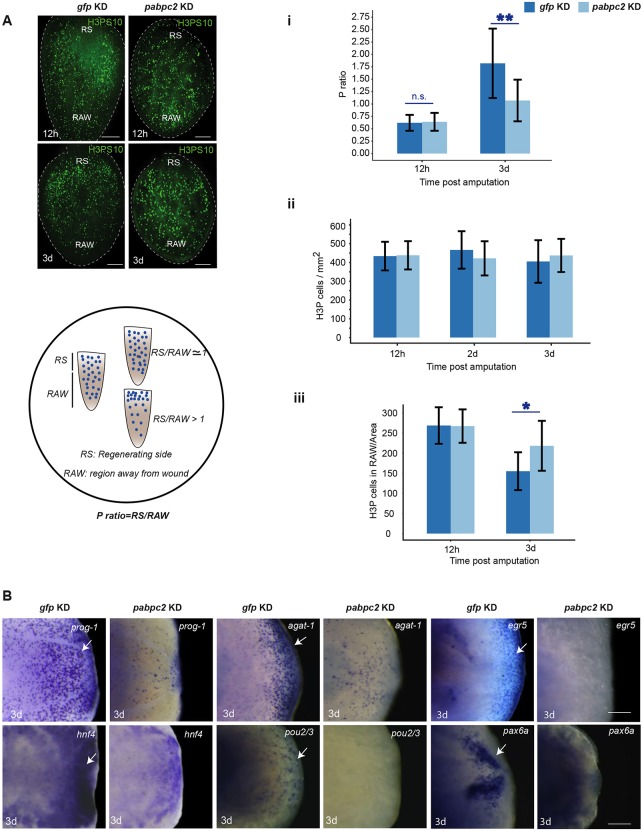
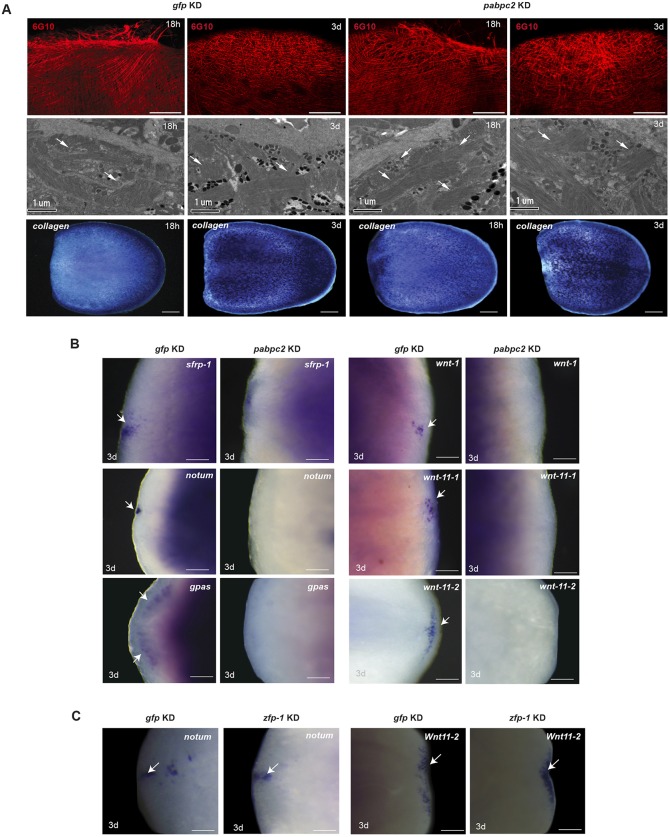
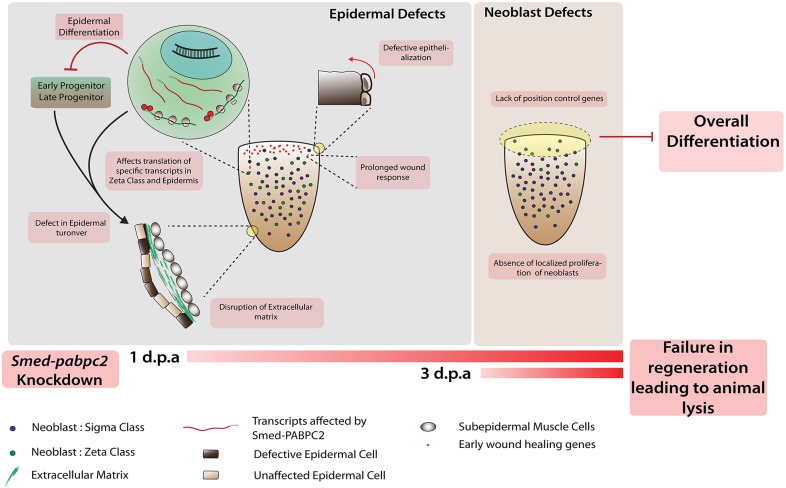
Identifying key cellular events that facilitate stem cell function and tissue organization is crucial for understanding the process of regeneration. Planarians are powerful model system to study regeneration and stem cell (neoblast) function. Here, using planaria, we show that the initial events of regeneration, such as epithelialization and epidermal organization are critically regulated by a novel cytoplasmic poly A-binding protein, SMED-PABPC2. Knockdown of smed-pabpc2 leads to defects in epidermal lineage specification, disorganization of epidermis and ECM, and deregulated wound healing, resulting in the selective failure of neoblast proliferation near the wound region. Polysome profiling suggests that epidermal lineage transcripts, including zfp-1, are translationally regulated by SMED-PABPC2. Together, our results uncover a novel role for SMED-PABPC2 in the maintenance of epidermal and ECM integrity, critical for wound healing and subsequent processes for regeneration.
Keywords: Epidermis; Neoblast; Planaria; Poly (A)-binding proteins; Regeneration.
© 2017. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
Similar articles
-
An insight into planarian regeneration.Cell Prolif. 2022 Sep;55(9):e13276. doi: 10.1111/cpr.13276. Epub 2022 Jul 10. Cell Prolif. 2022. PMID: 35811385 Free PMC article. Review.
-
The Akt signaling pathway is required for tissue maintenance and regeneration in planarians.BMC Dev Biol. 2016 Apr 11;16:7. doi: 10.1186/s12861-016-0107-z. BMC Dev Biol. 2016. PMID: 27068018 Free PMC article.
-
Smed-SmB, a member of the LSm protein superfamily, is essential for chromatoid body organization and planarian stem cell proliferation.Development. 2010 Apr;137(7):1055-65. doi: 10.1242/dev.042564. Development. 2010. PMID: 20215344
-
Epidermal turnover in the planarian Schmidtea mediterranea involves basal cell extrusion and intestinal digestion.Cell Rep. 2024 Jul 23;43(7):114305. doi: 10.1016/j.celrep.2024.114305. Epub 2024 Jun 20. Cell Rep. 2024. PMID: 38906148
-
Decoding Stem Cells: An Overview on Planarian Stem Cell Heterogeneity and Lineage Progression.Biomolecules. 2021 Oct 17;11(10):1532. doi: 10.3390/biom11101532. Biomolecules. 2021. PMID: 34680165 Free PMC article. Review.
Cited by
-
MRLC controls apoptotic cell death and functions to regulate epidermal development during planarian regeneration and homeostasis.Cell Prolif. 2024 Jan;57(1):e13524. doi: 10.1111/cpr.13524. Epub 2023 Jun 25. Cell Prolif. 2024. PMID: 37357415 Free PMC article.
-
The exon junction complex is required for stem and progenitor cell maintenance in planarians.Dev Biol. 2020 Jan 1;457(1):119-127. doi: 10.1016/j.ydbio.2019.09.010. Epub 2019 Sep 23. Dev Biol. 2020. PMID: 31557470 Free PMC article.
-
An insight into planarian regeneration.Cell Prolif. 2022 Sep;55(9):e13276. doi: 10.1111/cpr.13276. Epub 2022 Jul 10. Cell Prolif. 2022. PMID: 35811385 Free PMC article. Review.
-
Identification of rare, transient post-mitotic cell states that are induced by injury and required for whole-body regeneration in Schmidtea mediterranea.Nat Cell Biol. 2021 Sep;23(9):939-952. doi: 10.1038/s41556-021-00734-6. Epub 2021 Sep 2. Nat Cell Biol. 2021. PMID: 34475533 Free PMC article.
-
Comprehensive annotation and characterization of planarian tRNA and tRNA-derived fragments (tRFs).RNA. 2021 Apr;27(4):477-495. doi: 10.1261/rna.077701.120. Epub 2021 Jan 14. RNA. 2021. PMID: 33446492 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
