

Impact of Type III Secretion Effectors and of Phenoxyacetamide Inhibitors of Type III Secretion on Abscess Formation in a Mouse Model of Pseudomonas aeruginosa Infection
- PMID: 28807906
- PMCID: PMC5655109
- DOI: 10.1128/AAC.01202-17
Impact of Type III Secretion Effectors and of Phenoxyacetamide Inhibitors of Type III Secretion on Abscess Formation in a Mouse Model of Pseudomonas aeruginosa Infection
Abstract
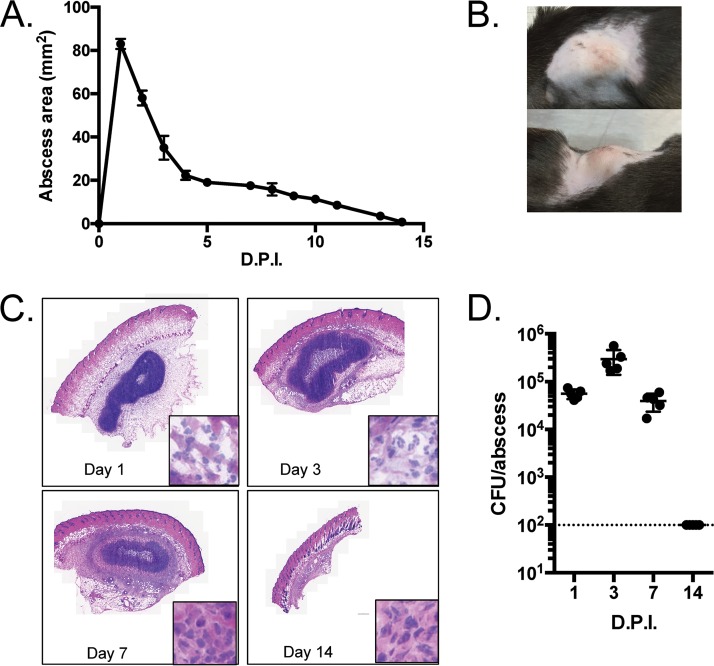
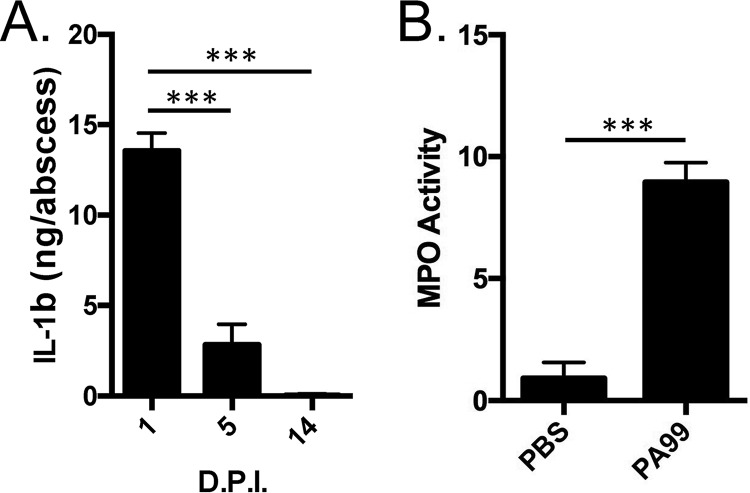
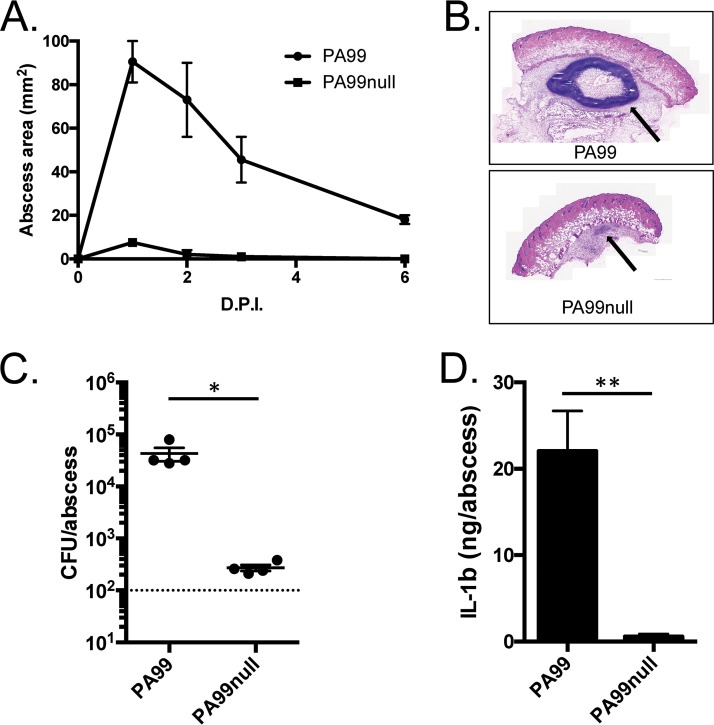
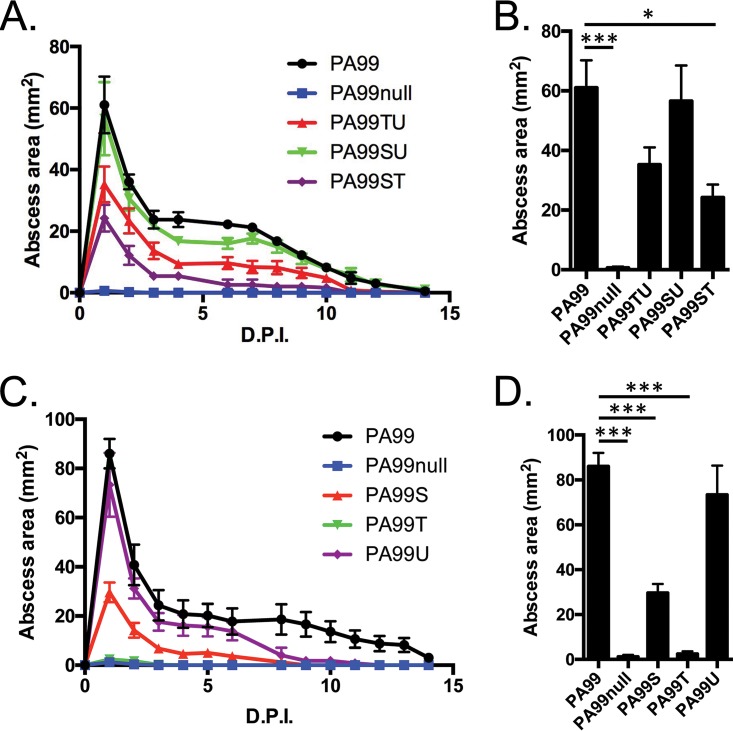
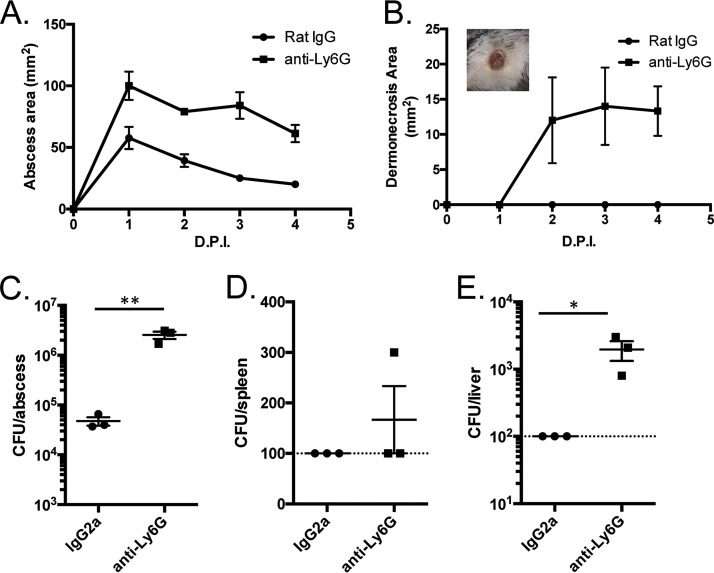
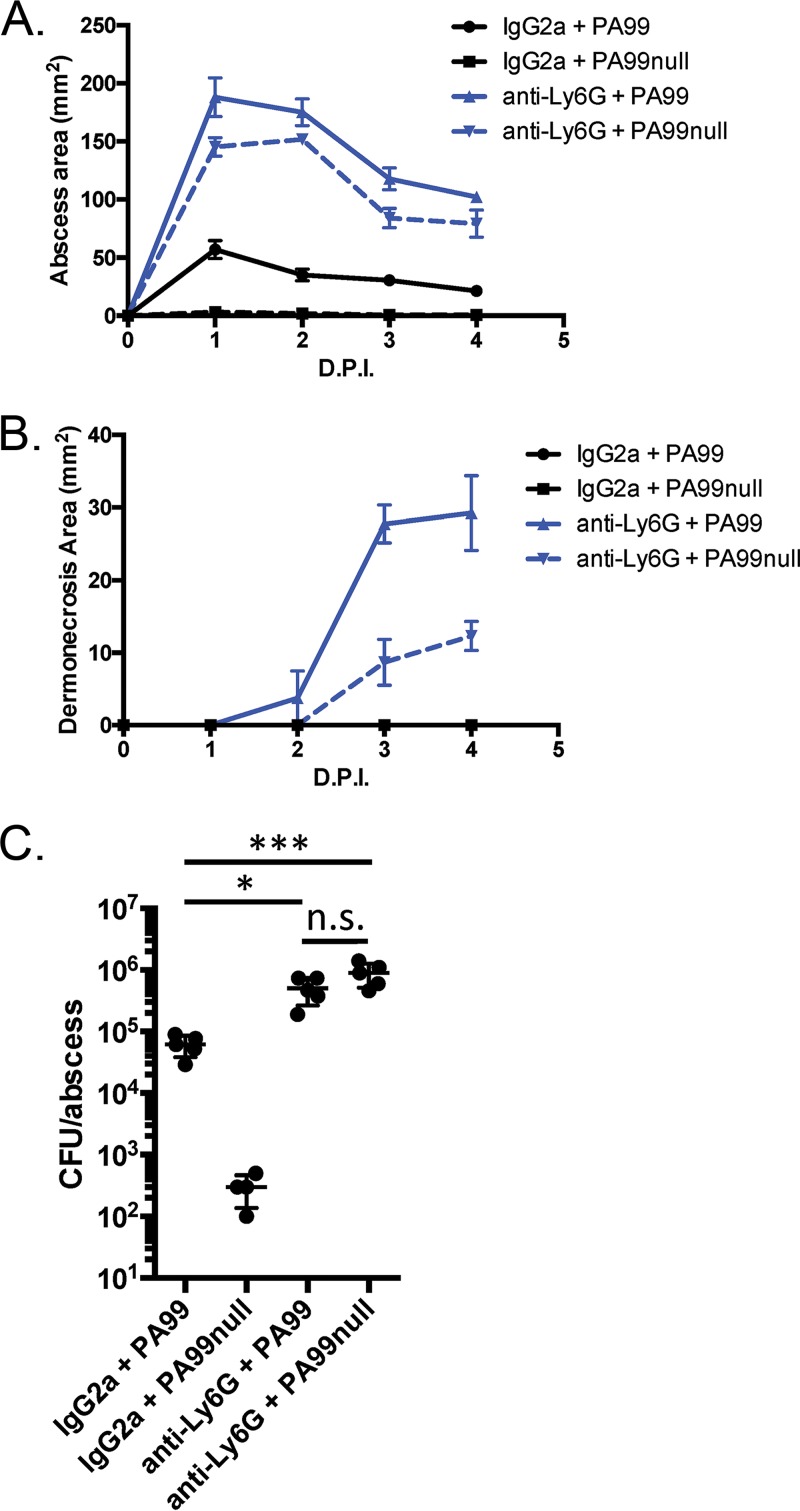
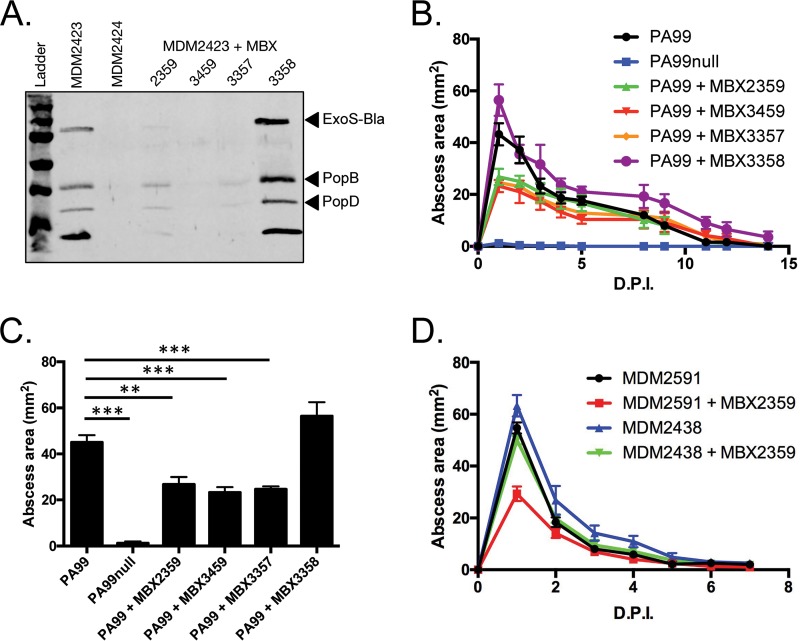
Pseudomonas aeruginosa is a leading cause of intra-abdominal infections, wound infections, and community-acquired folliculitis, each of which may involve macro- or microabscess formation. The rising incidence of multidrug resistance among P. aeruginosa isolates has increased both the economic burden and the morbidity and mortality associated with P. aeruginosa disease and necessitates a search for novel therapeutics. Previous work from our group detailed novel phenoxyacetamide inhibitors that block type III secretion and injection into host cells in vitro In this study, we used a mouse model of P. aeruginosa abscess formation to test the in vivo efficacy of these compounds against the P. aeruginosa type III secretion system (T3SS). Bacteria used the T3SS to intoxicate infiltrating neutrophils to establish abscesses. Despite this antagonism, sufficient numbers of functioning neutrophils remained for proper containment of the abscesses, as neutrophil depletion resulted in an increased abscess size, the formation of dermonecrotic lesions on the skin, and the dissemination of P. aeruginosa to internal organs. Consistent with the specificity of the T3SS-neutrophil interaction, P. aeruginosa bacteria lacking a functional T3SS were fully capable of causing abscesses in a neutropenic host. Phenoxyacetamide inhibitors attenuated abscess formation and aided in the immune clearance of the bacteria. Finally, a P. aeruginosa strain resistant to the phenoxyacetamide compound was fully capable of causing abscess formation even in the presence of the T3SS inhibitors. Together, our results further define the role of type III secretion in murine abscess formation and demonstrate the in vivo efficacy of phenoxyacetamide inhibitors in P. aeruginosa infection.
Keywords: Pseudomonas aeruginosa; abscesses; neutrophils; phenoxyacetamide inhibitors; type III secretion.
Copyright © 2017 American Society for Microbiology.
Figures

References
-
- Stryjewski M, Sexton DJ. 2003. Pseudomonas aeruginosa infections in specific types of patients and clinical settings, p 1–15. In Hauser A, Rello J (ed), Severe infections caused by Pseudomonas aeruginosa. Kluwer Academic Publishers, Boston, MA.
-
- Solomkin JS, Mazuski JE, Bradley JS, Rodvold KA, Goldstein EJ, Baron EJ, O'Neill PJ, Chow AW, Dellinger EP, Eachempati SR, Gorbach S, Hilfiker M, May AK, Nathens AB, Sawyer RG, Bartlett JG. 2010. Diagnosis and management of complicated intra-abdominal infection in adults and children: guidelines by the Surgical Infection Society and the Infectious Diseases Society of America. Clin Infect Dis 50:133–164. doi: 10.1086/649554. - DOI - PubMed
-
- Sievert DM, Ricks P, Edwards JR, Schneider A, Patel J, Srinivasan A, Kallen A, Limbago B, Fridkin S, National Healthcare Safety Network (NHSN) Team and Participating NHSN Facilities. 2013. Antimicrobial-resistant pathogens associated with healthcare-associated infections: summary of data reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2009-2010. Infect Control Hosp Epidemiol 34:1–14. doi: 10.1086/668770. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
