

Inhibition of Calcineurin or IMP Dehydrogenase Exerts Moderate to Potent Antiviral Activity against Norovirus Replication
- PMID: 28807916
- PMCID: PMC5655111
- DOI: 10.1128/AAC.01095-17
Inhibition of Calcineurin or IMP Dehydrogenase Exerts Moderate to Potent Antiviral Activity against Norovirus Replication
Abstract
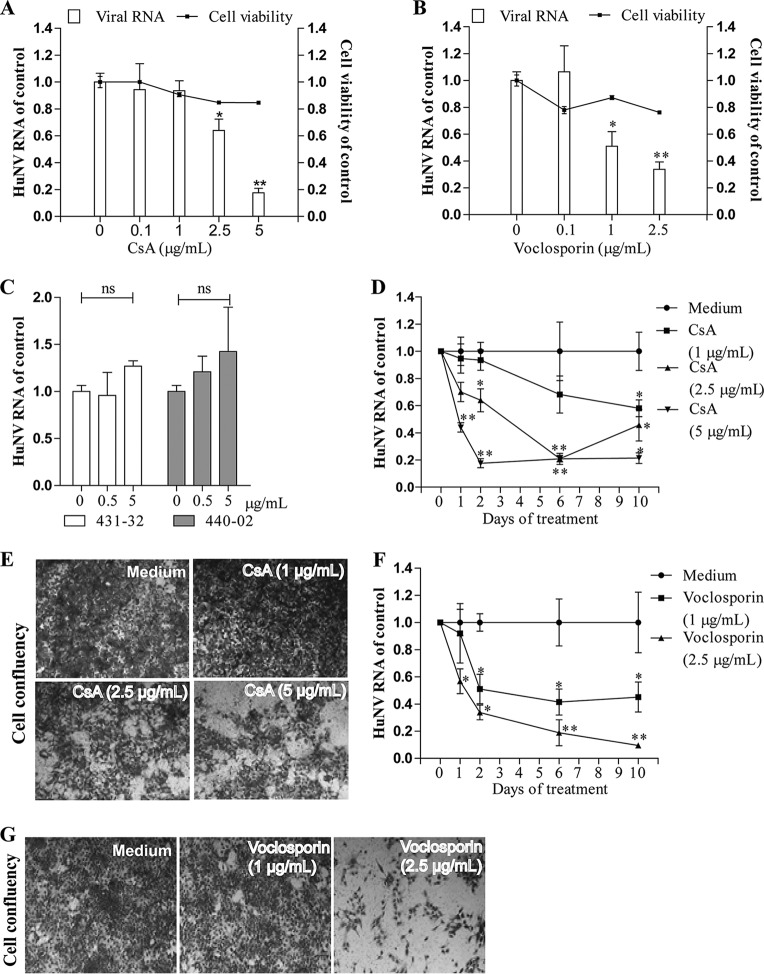
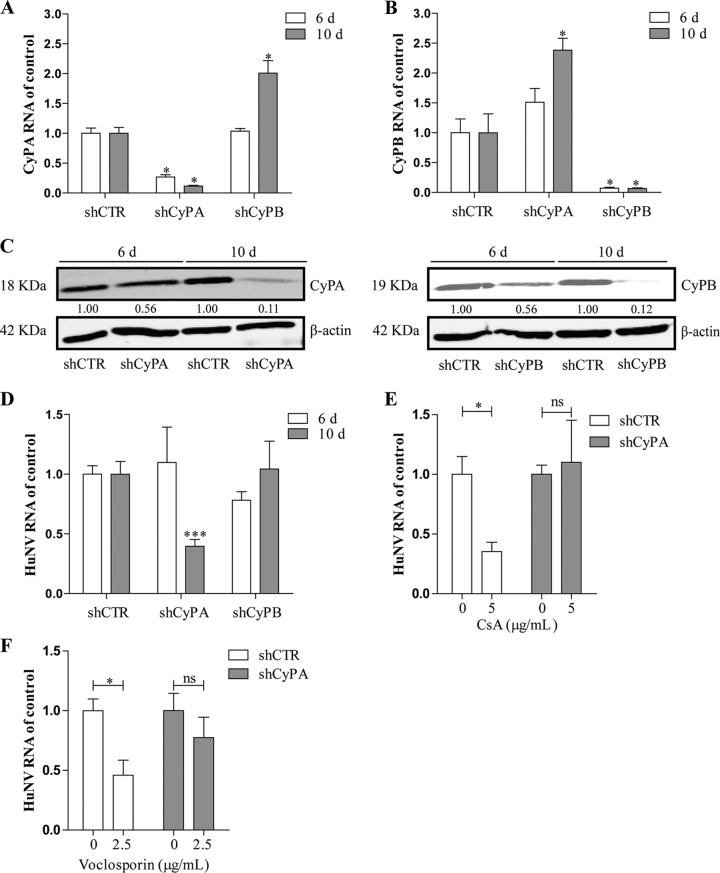
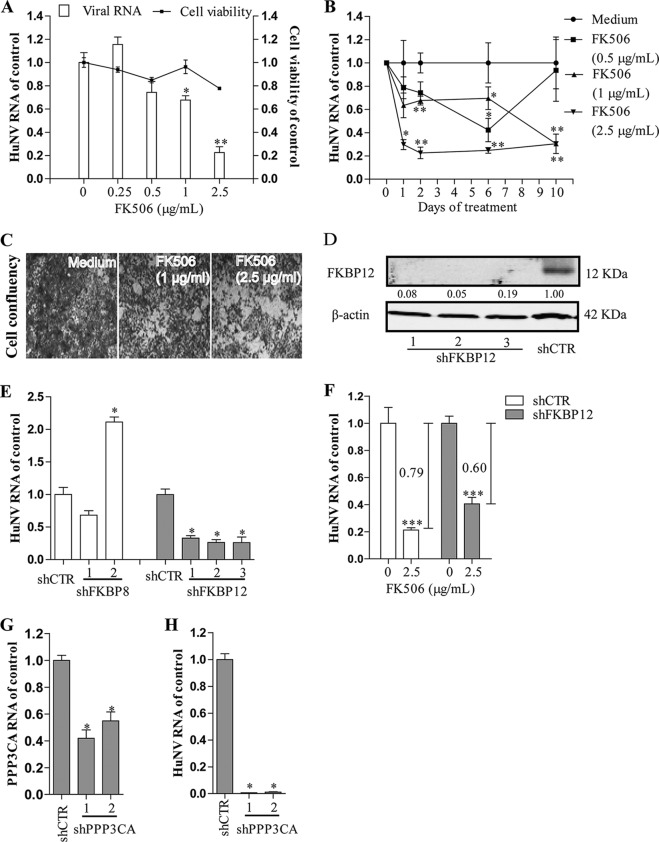
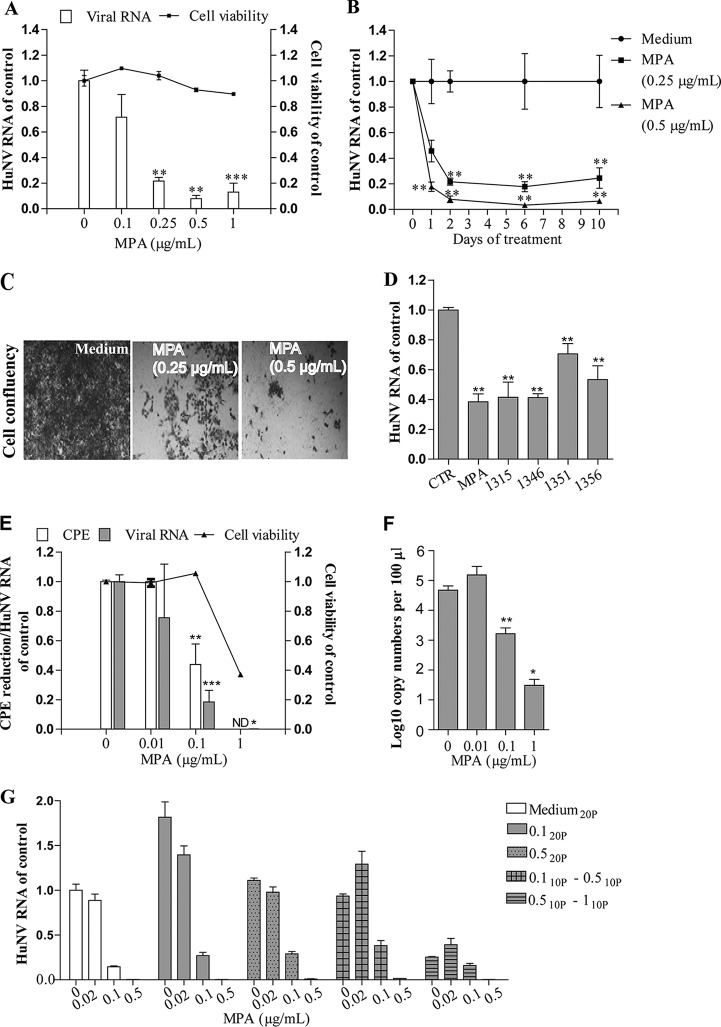
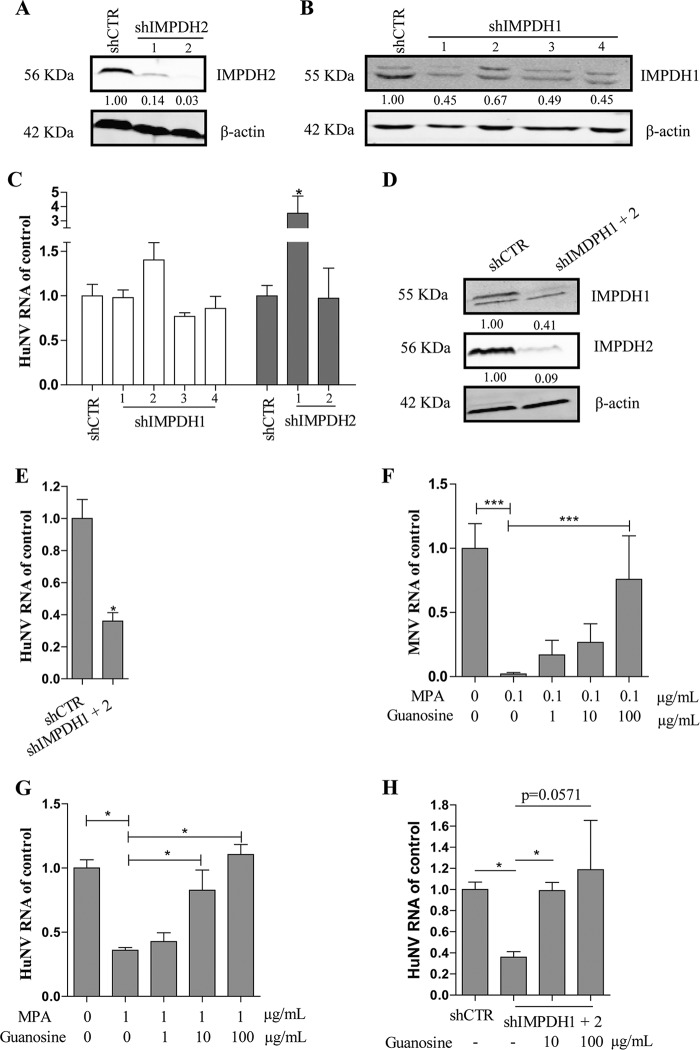
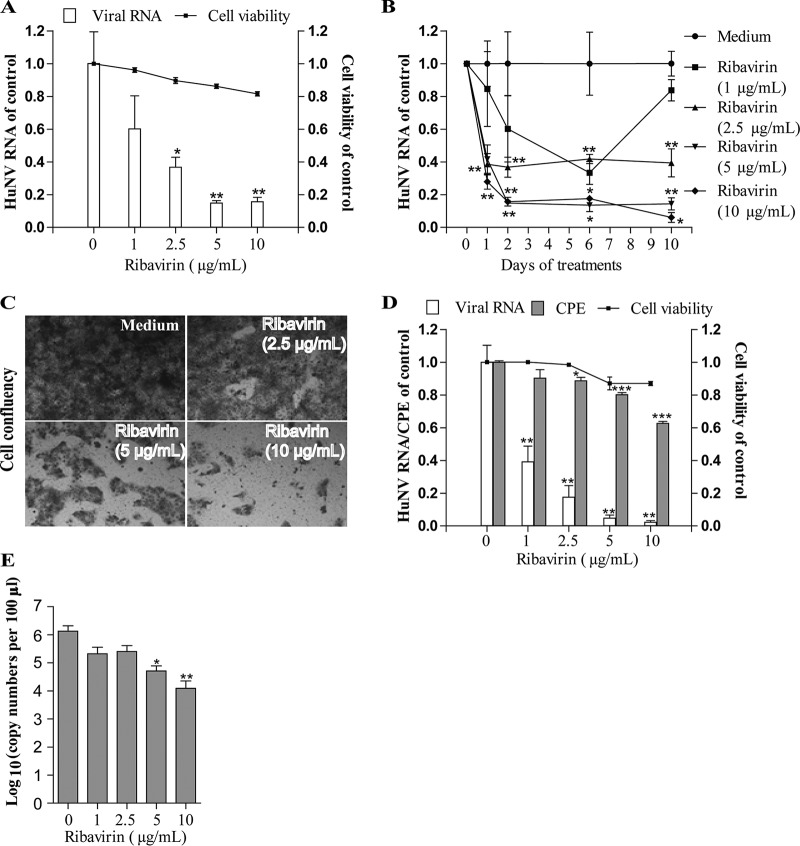
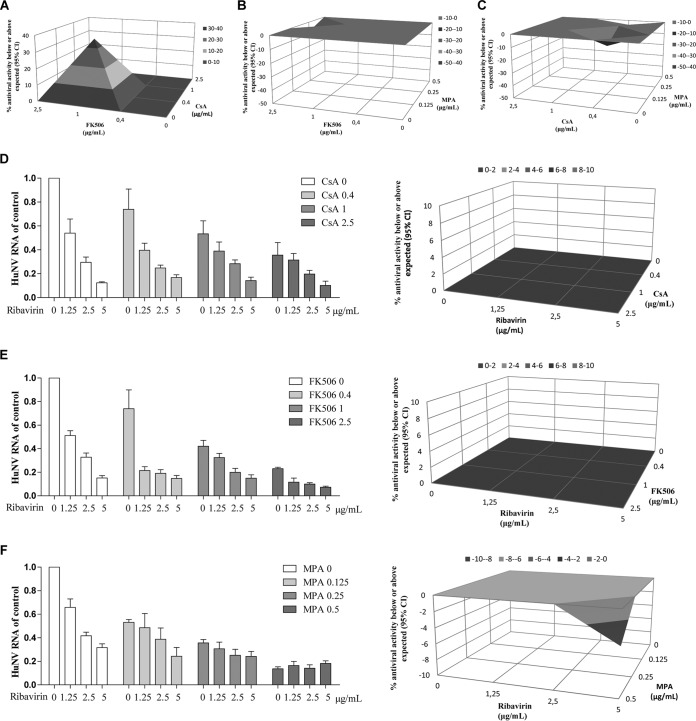
Norovirus is a major cause of acute gastroenteritis worldwide and has emerged as an important issue of chronic infection in transplantation patients. Since no approved antiviral is available, we evaluated the effects of different immunosuppressants and ribavirin on norovirus and explored their mechanisms of action by using a human norovirus (HuNV) replicon-harboring model and a surrogate murine norovirus (MNV) infectious model. The roles of the corresponding drug targets were investigated by gain- or loss-of-function approaches. We found that the calcineurin inhibitors cyclosporine (CsA) and tacrolimus (FK506) moderately inhibited HuNV replication. Gene silencing of their cellular targets, cyclophilin A, FKBP12, and calcineurin, significantly inhibited HuNV replication. A low concentration, therapeutically speaking, of mycophenolic acid (MPA), an uncompetitive IMP dehydrogenase (IMPDH) inhibitor, potently and rapidly inhibited norovirus replication and ultimately cleared HuNV replicons without inducible resistance following long-term drug exposure. Knockdown of the MPA cellular targets IMPDH1 and IMPDH2 suppressed HuNV replication. Consistent with the nucleotide-synthesizing function of IMPDH, exogenous guanosine counteracted the antinorovirus effects of MPA. Furthermore, the competitive IMPDH inhibitor ribavirin efficiently inhibited norovirus and resulted in an additive effect when combined with immunosuppressants. The results from this study demonstrate that calcineurin phosphatase activity and IMPDH guanine synthase activity are crucial in sustaining norovirus infection; thus, they can be therapeutically targeted. Our results suggest that MPA shall be preferentially considered immunosuppressive medication for transplantation patients at risk of norovirus infection, whereas ribavirin represents as a potential antiviral for both immunocompromised and immunocompetent patients with norovirus gastroenteritis.
Keywords: calcineurin inhibitors; cell culture; cell culture model; mycophenolic acid; norovirus; noroviruses; ribavirin.
Copyright © 2017 American Society for Microbiology.
Figures
References
-
- McAtee CL, Webman R, Gilman RH, Mejia C, Bern C, Apaza S, Espetia S, Pajuelo M, Saito M, Challappa R, Soria R, Ribera JP, Lozano D, Torrico F. 2016. Burden of norovirus and rotavirus in children after rotavirus vaccine introduction, Cochabamba, Bolivia. Am J Trop Med Hyg 94:212–217. doi: 10.4269/ajtmh.15-0203. - DOI - PMC - PubMed
-
- Hoa Tran TN, Trainor E, Nakagomi T, Cunliffe NA, Nakagomi O. 2013. Molecular epidemiology of noroviruses associated with acute sporadic gastroenteritis in children: global distribution of genogroups, genotypes and GII.4 variants. J Clin Virol 56:185–193. doi: 10.1016/j.jcv.2012.11.011. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
