

Human mucosal-associated invariant T (MAIT) cells possess capacity for B cell help
- PMID: 28807929
- PMCID: PMC5636046
- DOI: 10.1189/jlb.4A0317-116R
Human mucosal-associated invariant T (MAIT) cells possess capacity for B cell help
Abstract
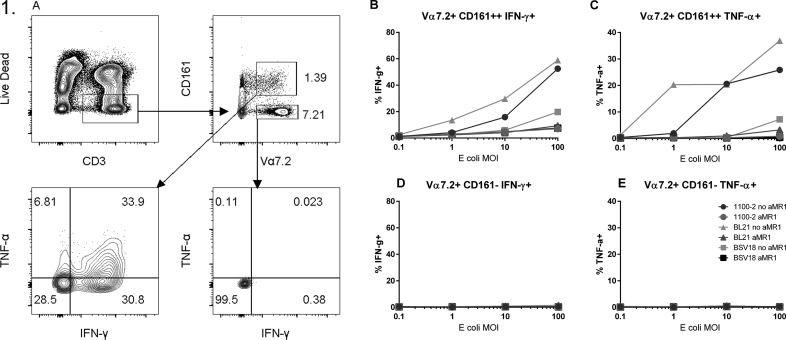
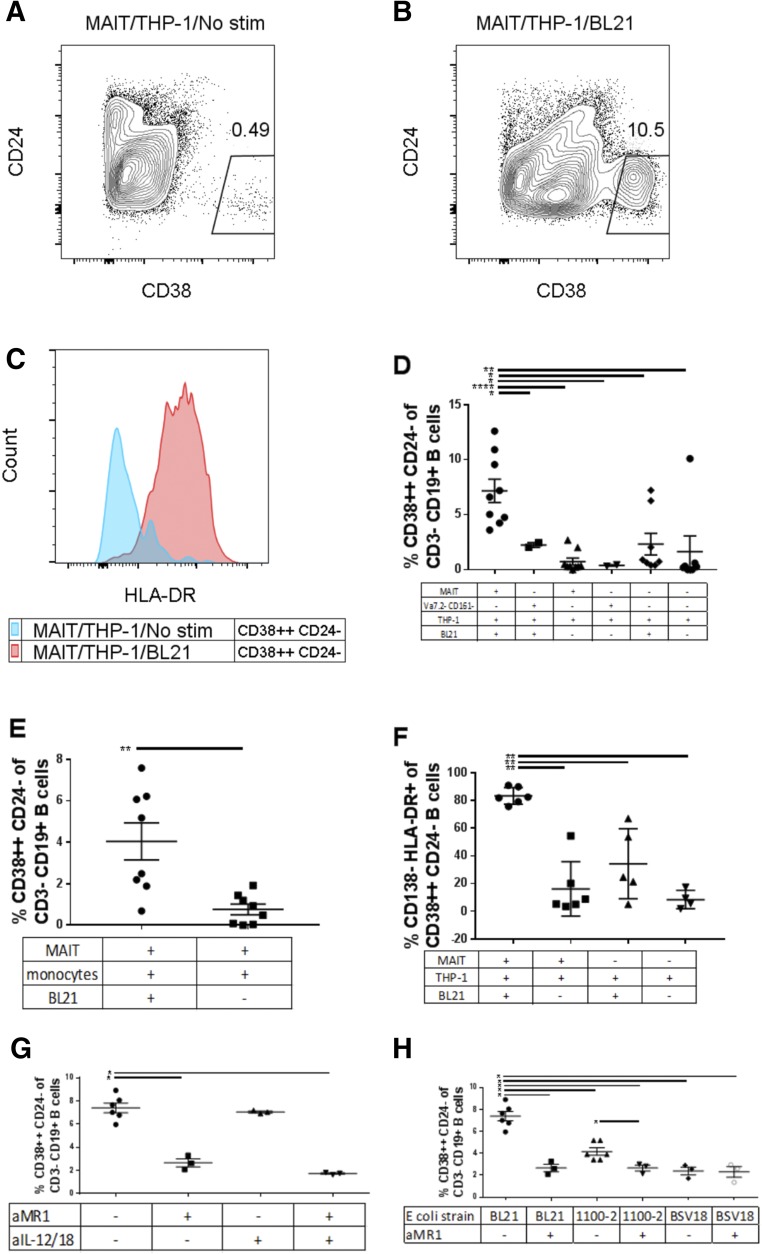
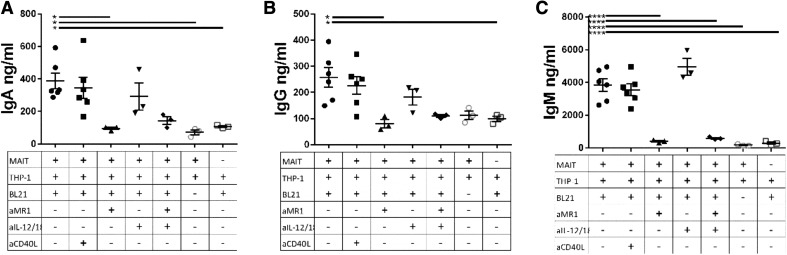
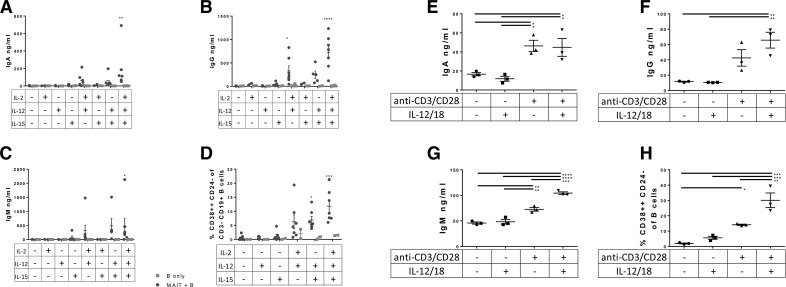
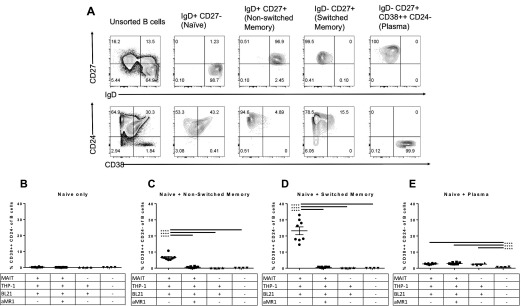
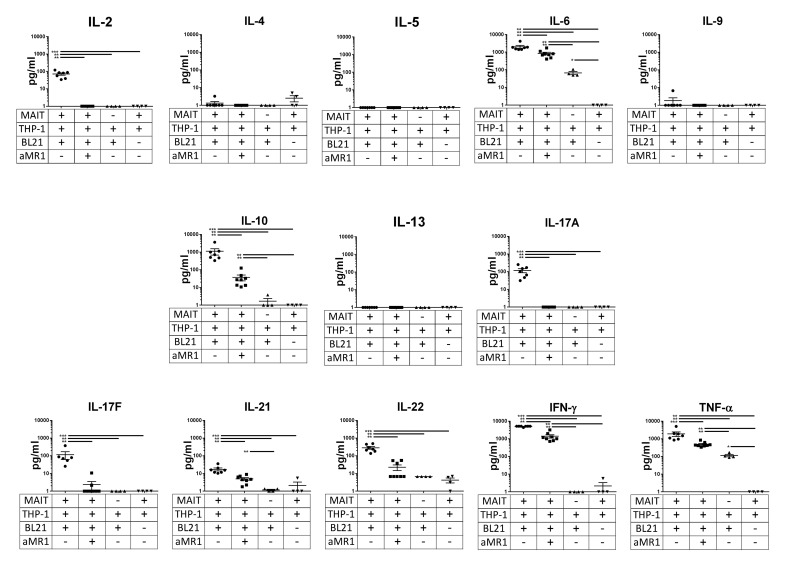
Mucosal-associated invariant T (MAIT) cells are an innate-like T cell subset, restricted by the nonclassic MHC class I-related protein MR1 and enriched at mucosal sites. Human studies have shown an association between MAIT cells and pathogen-specific antibody responses. In this study, we investigate the effect of human MAIT cells on B cells ex vivo. We found that supernatants from microbe- or cytokine-stimulated MAIT cells, when added to purified autologous B cells, increase frequencies of plasmablasts and promote IgA, IgG, and IgM production. We found effects to be mostly MR1-dependent and that the increases in plasmablasts are likely a result of increased differentiation from memory B cells. Furthermore, microbe-activated MAIT cell supernatant contains multiple cytokines known to stimulate B cells, including IL-6, -10, and -21. This study thus provides the first direct evidence of a newly identified role of MAIT cells in providing help to B cells.
Keywords: B cell help; MR-1; antibody; plasmablast.
© Society for Leukocyte Biology.
Figures
References
-
- Dellabona P., Abrignani S., Casorati G. (2014) iNKT-cell help to B cells: a cooperative job between innate and adaptive immune responses. Eur. J. Immunol. 44, 2230–2237. - PubMed
-
- Kjer-Nielsen L., Patel O., Corbett A. J., Le Nours J., Meehan B., Liu L., Bhati M., Chen Z., Kostenko L., Reantragoon R., Williamson N. A., Purcell A. W., Dudek N. L., McConville M. J., O’Hair R. A., Khairallah G. N., Godfrey D. I., Fairlie D. P., Rossjohn J., McCluskey J. (2012) MR1 presents microbial vitamin B metabolites to MAIT cells. Nature 491, 717–723. - PubMed
-
- Walker L. J., Kang Y. H., Smith M. O., Tharmalingham H., Ramamurthy N., Fleming V. M., Sahgal N., Leslie A., Oo Y., Geremia A., Scriba T. J., Hanekom W. A., Lauer G. M., Lantz O., Adams D. H., Powrie F., Barnes E., Klenerman P. (2012) Human MAIT and CD8αα cells develop from a pool of type-17 precommitted CD8+ T cells. Blood 119, 422–433. - PMC - PubMed
-
- Eckle S. B., Birkinshaw R. W., Kostenko L., Corbett A. J., McWilliam H. E., Reantragoon R., Chen Z., Gherardin N. A., Beddoe T., Liu L., Patel O., Meehan B., Fairlie D. P., Villadangos J. A., Godfrey D. I., Kjer-Nielsen L., McCluskey J., Rossjohn J. (2014) A molecular basis underpinning the T cell receptor heterogeneity of mucosal-associated invariant T cells. J. Exp. Med. 211, 1585–1600. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
