

Immune evasion by a staphylococcal inhibitor of myeloperoxidase
- PMID: 28808028
- PMCID: PMC5584439
- DOI: 10.1073/pnas.1707032114
Immune evasion by a staphylococcal inhibitor of myeloperoxidase
Abstract
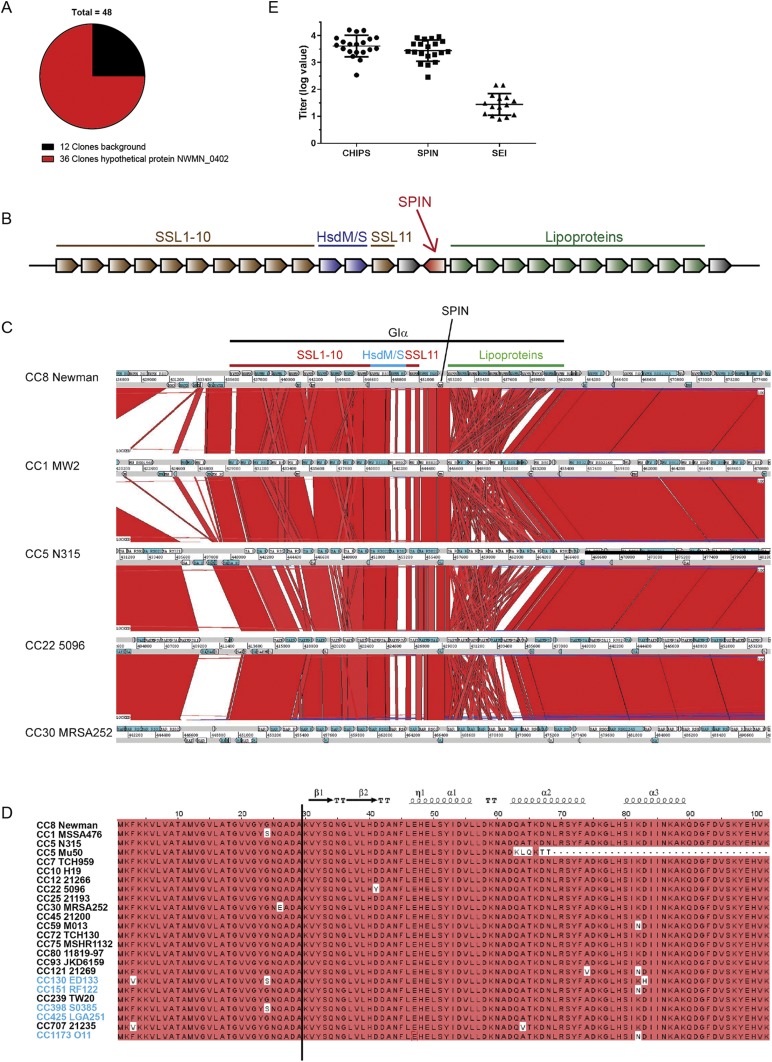
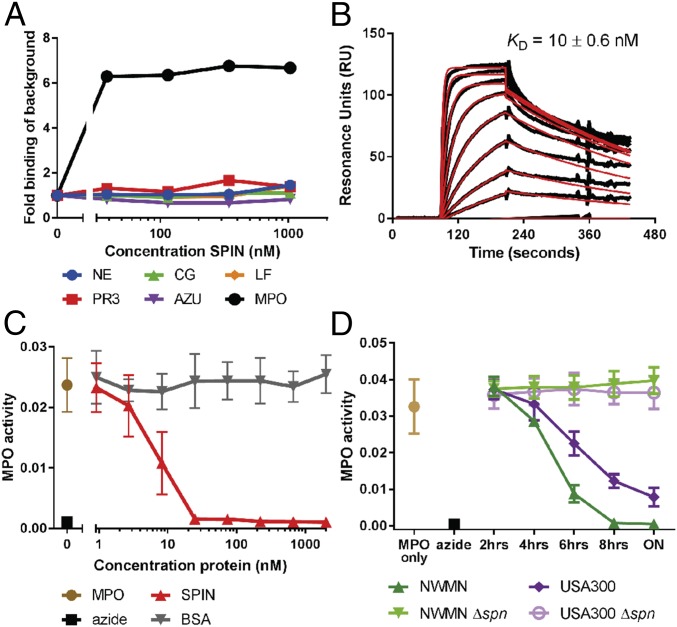
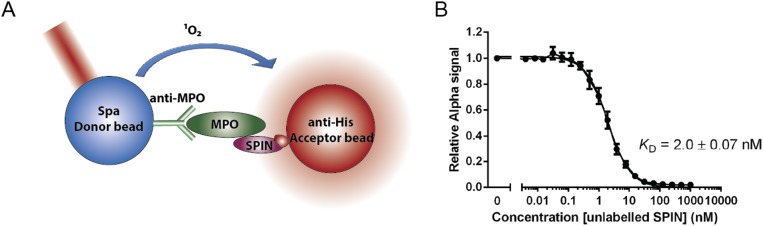
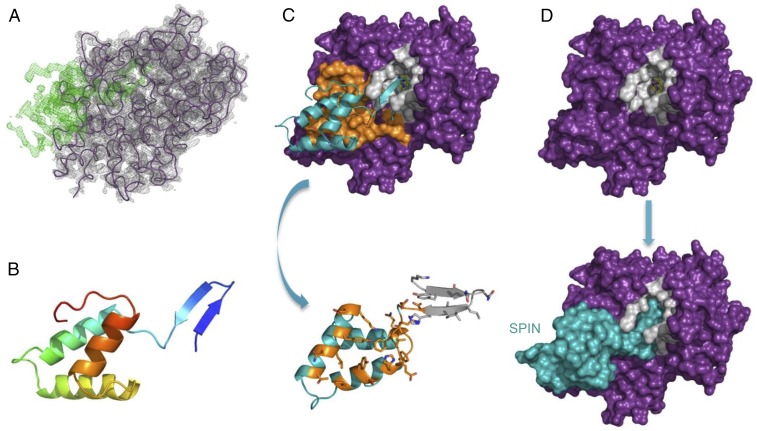
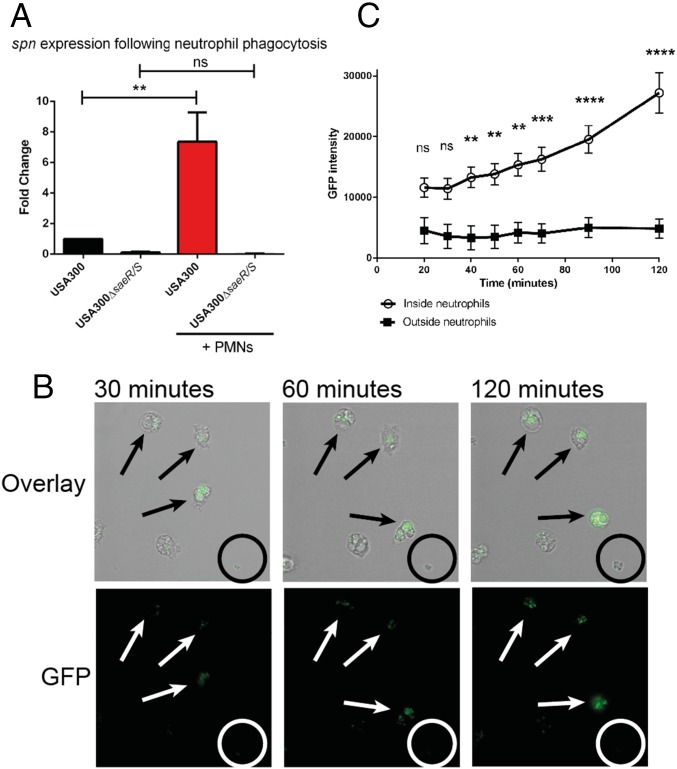
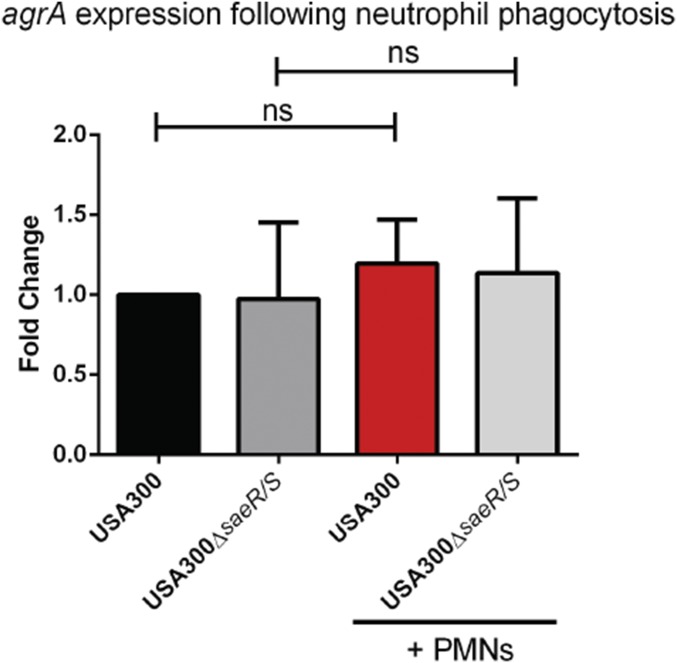
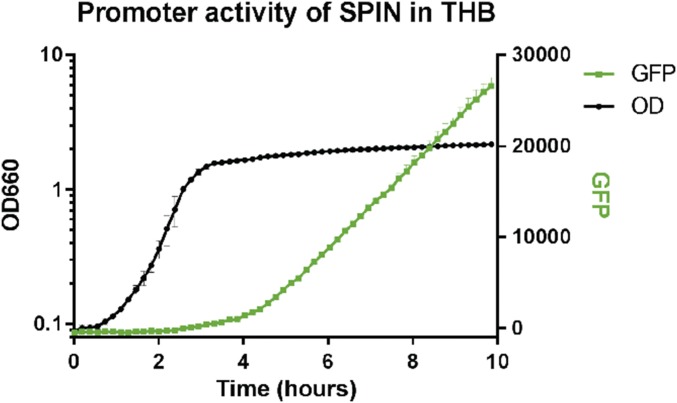
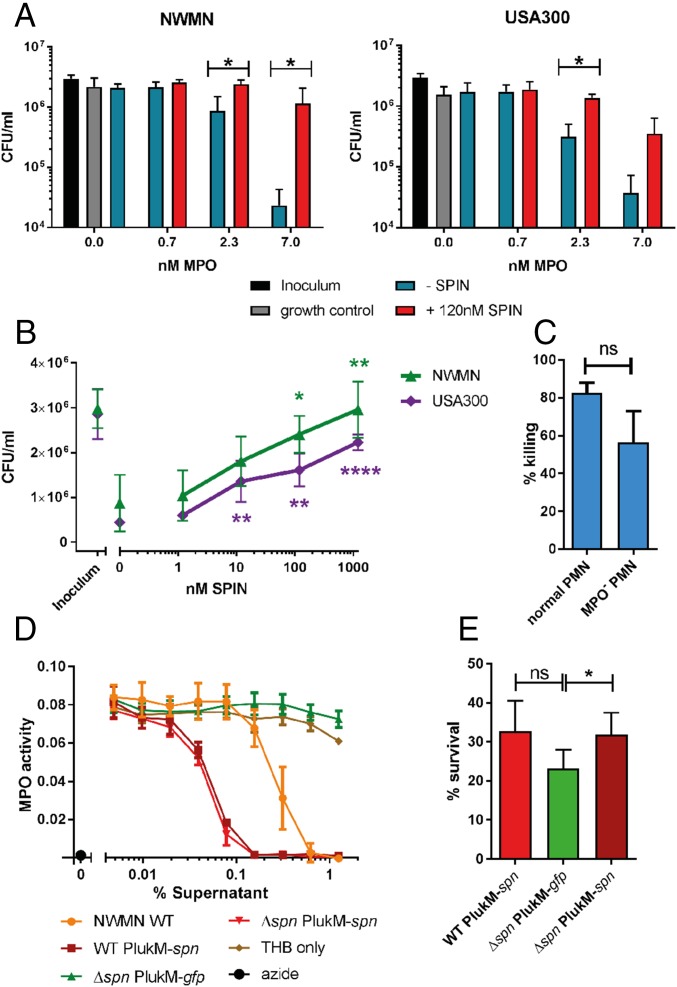
Staphylococcus aureus is highly adapted to its host and has evolved many strategies to resist opsonization and phagocytosis. Even after uptake by neutrophils, S. aureus shows resistance to killing, which suggests the presence of phagosomal immune evasion molecules. With the aid of secretome phage display, we identified a highly conserved protein that specifically binds and inhibits human myeloperoxidase (MPO), a major player in the oxidative defense of neutrophils. We have named this protein "staphylococcal peroxidase inhibitor" (SPIN). To gain insight into inhibition of MPO by SPIN, we solved the cocrystal structure of SPIN bound to a recombinant form of human MPO at 2.4-Å resolution. This structure reveals that SPIN acts as a molecular plug that prevents H2O2 substrate access to the MPO active site. In subsequent experiments, we observed that SPIN expression increases inside the neutrophil phagosome, where MPO is located, compared with outside the neutrophil. Moreover, bacteria with a deleted gene encoding SPIN showed decreased survival compared with WT bacteria after phagocytosis by neutrophils. Taken together, our results demonstrate that S. aureus secretes a unique proteinaceous MPO inhibitor to enhance survival by interfering with MPO-mediated killing.
Keywords: Staphylococcus aureus; immune evasion; myeloperoxidase; neutrophil; phagocytosis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

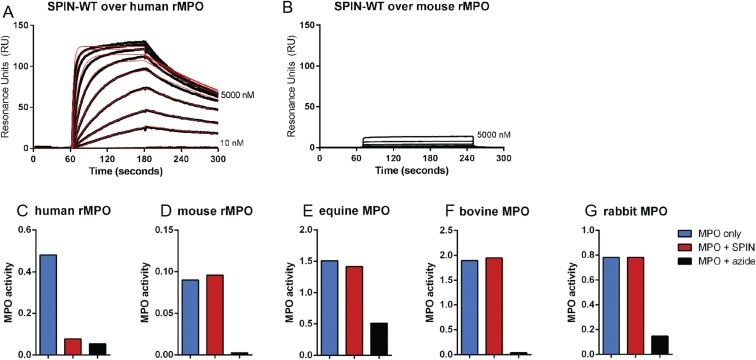
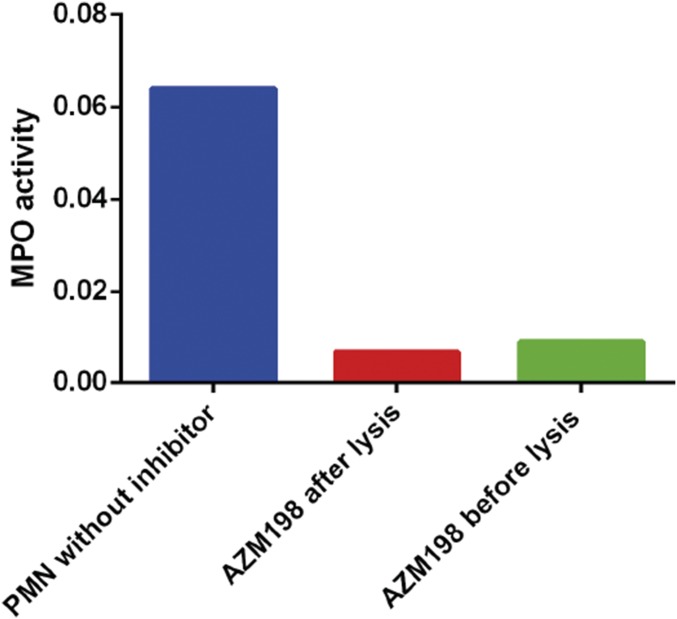
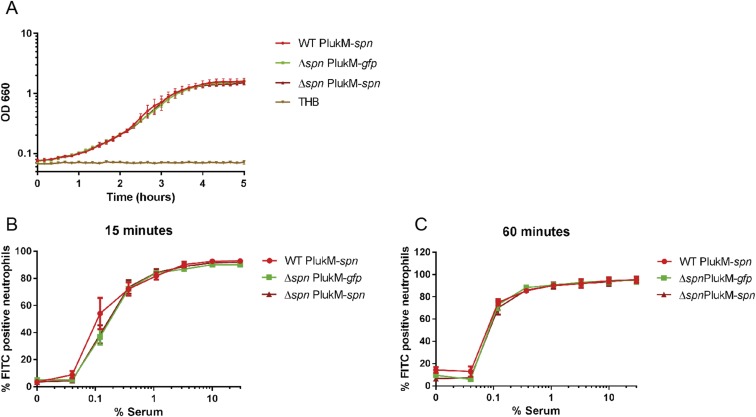
References
-
- Lowy FD. Staphylococcus aureus infections. N Engl J Med. 1998;339:520–532. - PubMed
-
- Faurschou M, Borregaard N. Neutrophil granules and secretory vesicles in inflammation. Microbes Infect. 2003;5:1317–1327. - PubMed
-
- Babior BM, Lambeth JD, Nauseef W. The neutrophil NADPH oxidase. Arch Biochem Biophys. 2002;397:342–344. - PubMed
-
- Klebanoff SJ, Hamon CB. Role of myeloperoxidase-mediated antimicrobial systems in intact leukocytes. J Reticuloendothel Soc. 1972;12:170–196. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
