

Rapid RNase L-driven arrest of protein synthesis in the dsRNA response without degradation of translation machinery
- PMID: 28808124
- PMCID: PMC5648034
- DOI: 10.1261/rna.062000.117
Rapid RNase L-driven arrest of protein synthesis in the dsRNA response without degradation of translation machinery
Abstract
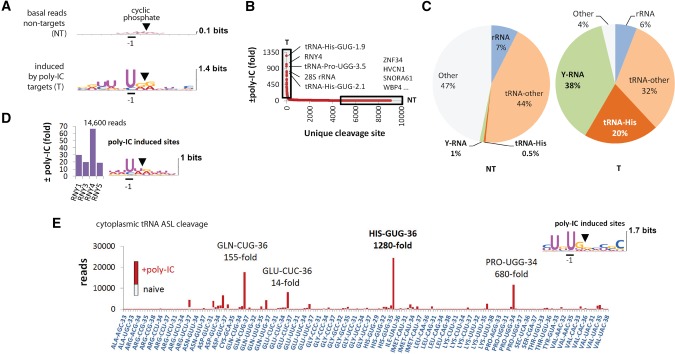
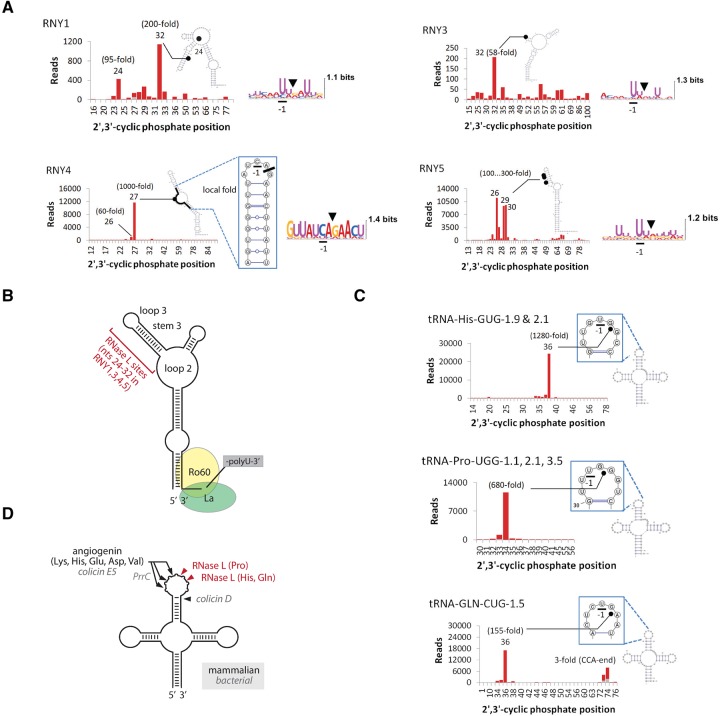
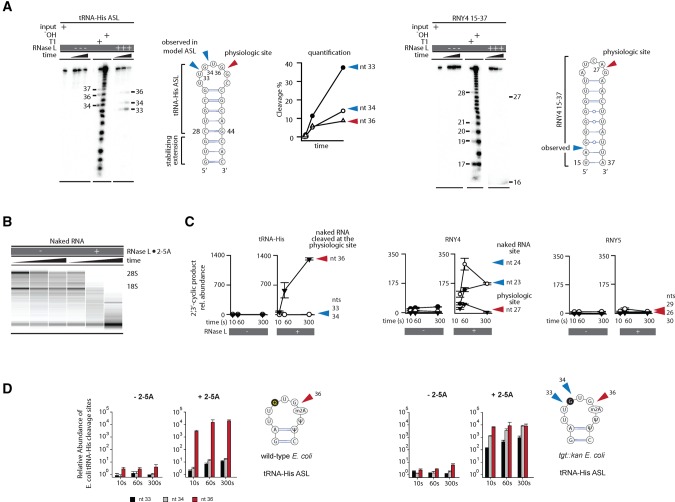
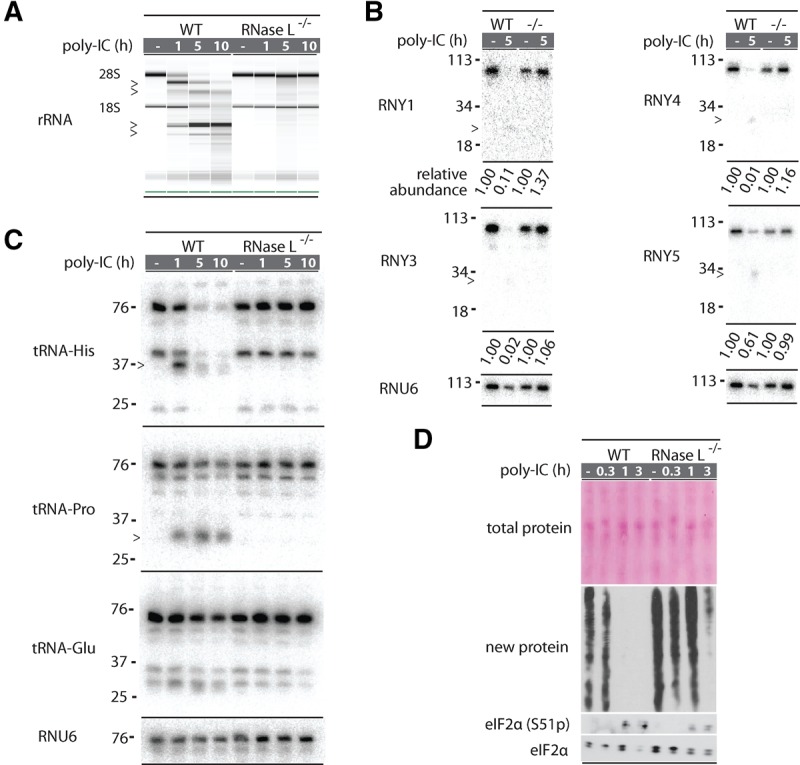
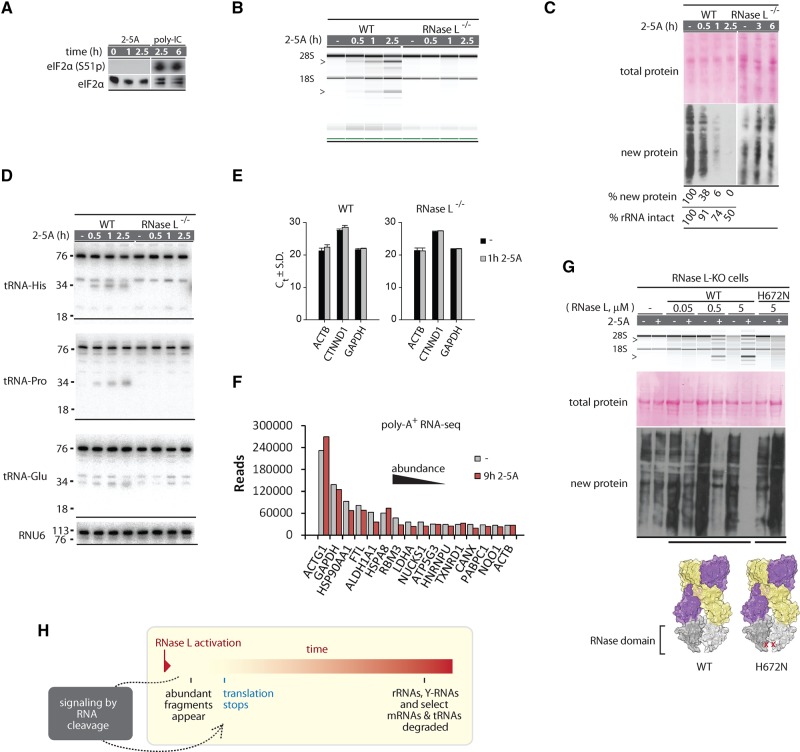
Mammalian cells respond to double-stranded RNA (dsRNA) by activating a translation-inhibiting endoribonuclease, RNase L. Consensus in the field indicates that RNase L arrests protein synthesis by degrading ribosomal RNAs (rRNAs) and messenger RNAs (mRNAs). However, here we provide evidence for a different and far more efficient mechanism. By sequencing abundant RNA fragments generated by RNase L in human cells, we identify site-specific cleavage of two groups of noncoding RNAs: Y-RNAs, whose function is poorly understood, and cytosolic tRNAs, which are essential for translation. Quantitative analysis of human RNA cleavage versus nascent protein synthesis in lung carcinoma cells shows that RNase L stops global translation when tRNAs, as well as rRNAs and mRNAs, are still intact. Therefore, RNase L does not have to degrade the translation machinery to stop protein synthesis. Our data point to a rapid mechanism that transforms a subtle RNA cleavage into a cell-wide translation arrest.
Keywords: RNase L; Y-RNA; signaling; tRNA; translation.
© 2017 Donovan et al.; Published by Cold Spring Harbor Laboratory Press for the RNA Society.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases