

Testing Convergent Evolution in Auditory Processing Genes between Echolocating Mammals and the Aye-Aye, a Percussive-Foraging Primate
- PMID: 28810710
- PMCID: PMC5553384
- DOI: 10.1093/gbe/evx140
Testing Convergent Evolution in Auditory Processing Genes between Echolocating Mammals and the Aye-Aye, a Percussive-Foraging Primate
Abstract
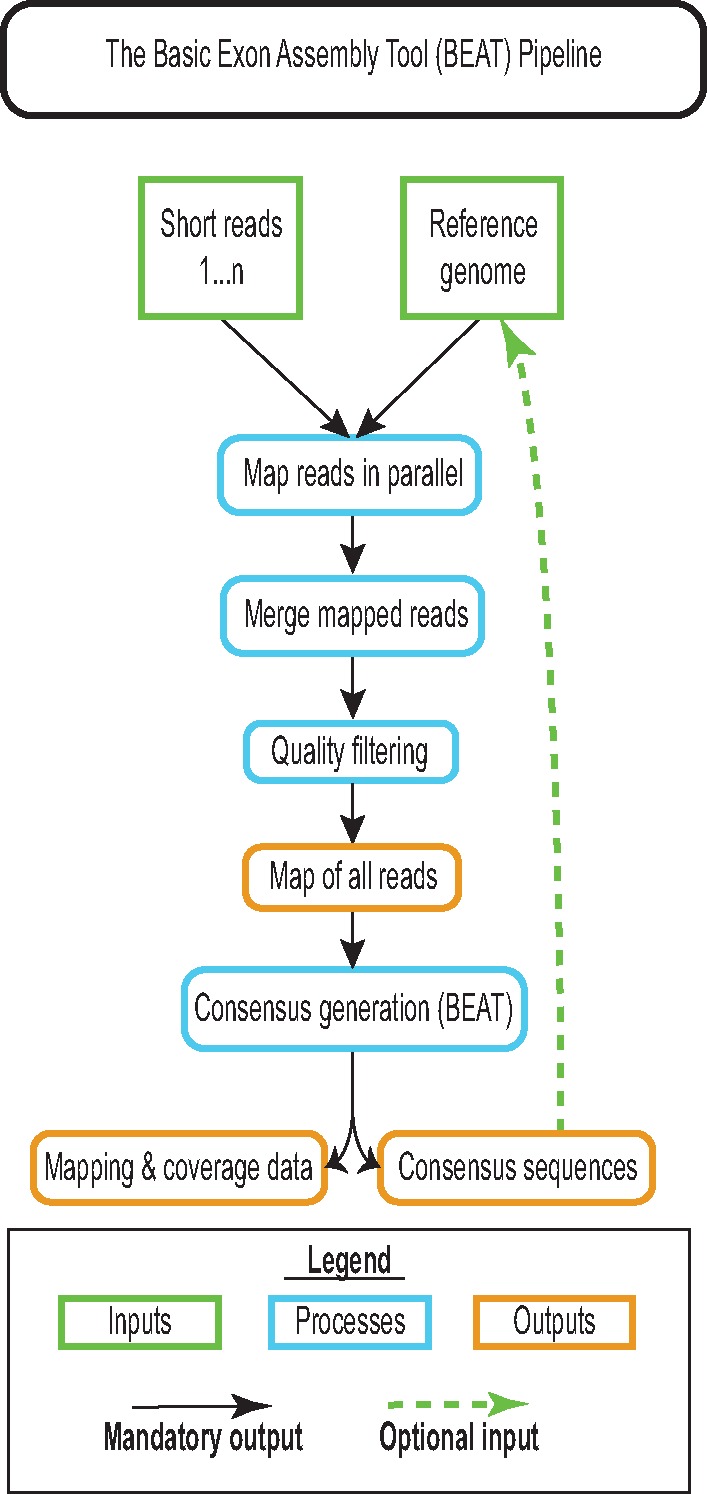
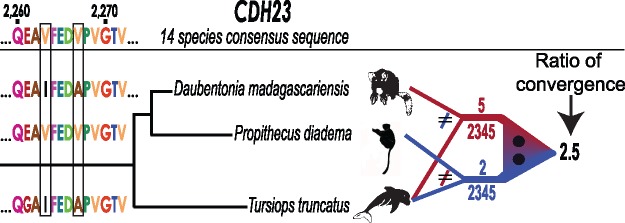
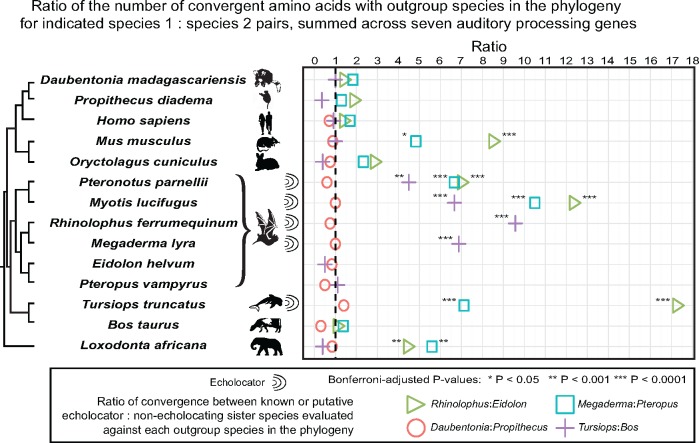
Several taxonomically distinct mammalian groups-certain microbats and cetaceans (e.g., dolphins)-share both morphological adaptations related to echolocation behavior and strong signatures of convergent evolution at the amino acid level across seven genes related to auditory processing. Aye-ayes (Daubentonia madagascariensis) are nocturnal lemurs with a specialized auditory processing system. Aye-ayes tap rapidly along the surfaces of trees, listening to reverberations to identify the mines of wood-boring insect larvae; this behavior has been hypothesized to functionally mimic echolocation. Here we investigated whether there are signals of convergence in auditory processing genes between aye-ayes and known mammalian echolocators. We developed a computational pipeline (Basic Exon Assembly Tool) that produces consensus sequences for regions of interest from shotgun genomic sequencing data for nonmodel organisms without requiring de novo genome assembly. We reconstructed complete coding region sequences for the seven convergent echolocating bat-dolphin genes for aye-ayes and another lemur. We compared sequences from these two lemurs in a phylogenetic framework with those of bat and dolphin echolocators and appropriate nonecholocating outgroups. Our analysis reaffirms the existence of amino acid convergence at these loci among echolocating bats and dolphins; some methods also detected signals of convergence between echolocating bats and both mice and elephants. However, we observed no significant signal of amino acid convergence between aye-ayes and echolocating bats and dolphins, suggesting that aye-aye tap-foraging auditory adaptations represent distinct evolutionary innovations. These results are also consistent with a developing consensus that convergent behavioral ecology does not reliably predict convergent molecular evolution.
Keywords: comparative genomics; convergent evolution; evolutionary ecology.
© The Author 2017. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures
References
-
- Au WWL, Simmons JA.. 2007. Echolocation in dolphins and bats. Phys Today [Internet] 60(9): 40–45.
-
- Cartmill M. 1974. Daubentonia, Dactylopsila, woodpeckers and klinorhynchy In: Martin RD, Doyle GA, Walker AC, editors. Prosimian biology. London: Duckworth; p. 655–670.
-
- Coleman MN, Ross CF.. 2004. Primate auditory diversity and its influence on hearing performance. Anat Rec A Discov Mol Cell Evol Biol. 281: 1123–1137. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
