

Human microcephaly protein RTTN interacts with STIL and is required to build full-length centrioles
- PMID: 28811500
- PMCID: PMC5558016
- DOI: 10.1038/s41467-017-00305-0
Human microcephaly protein RTTN interacts with STIL and is required to build full-length centrioles
Abstract
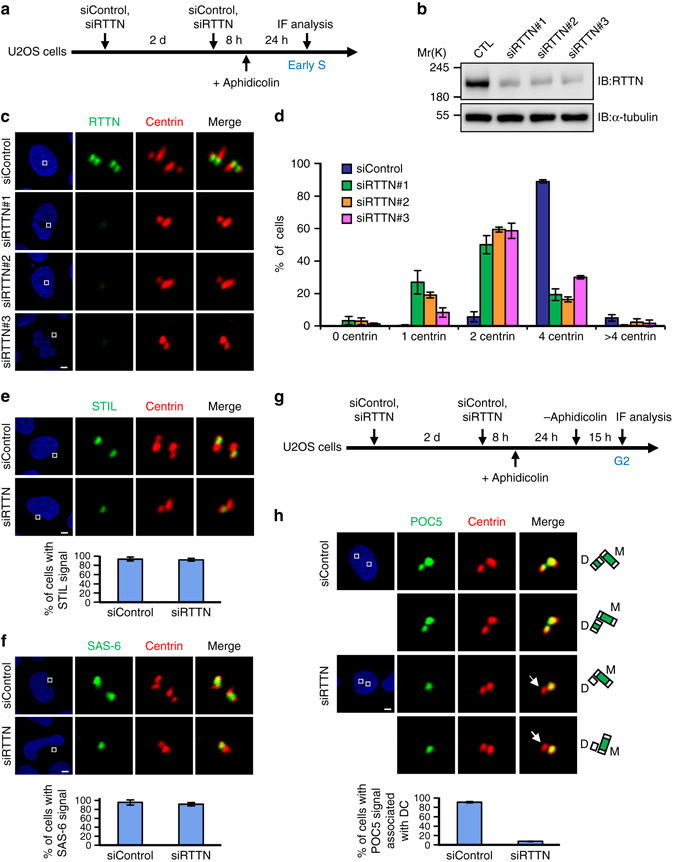
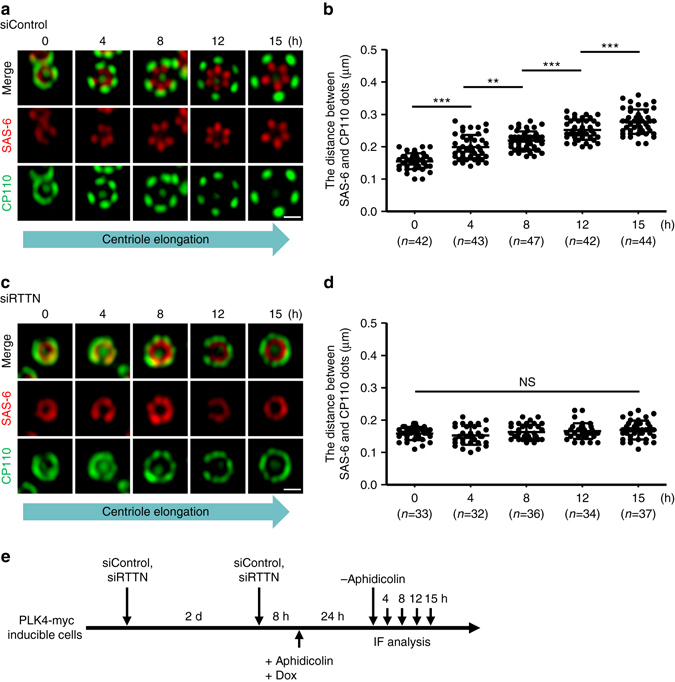
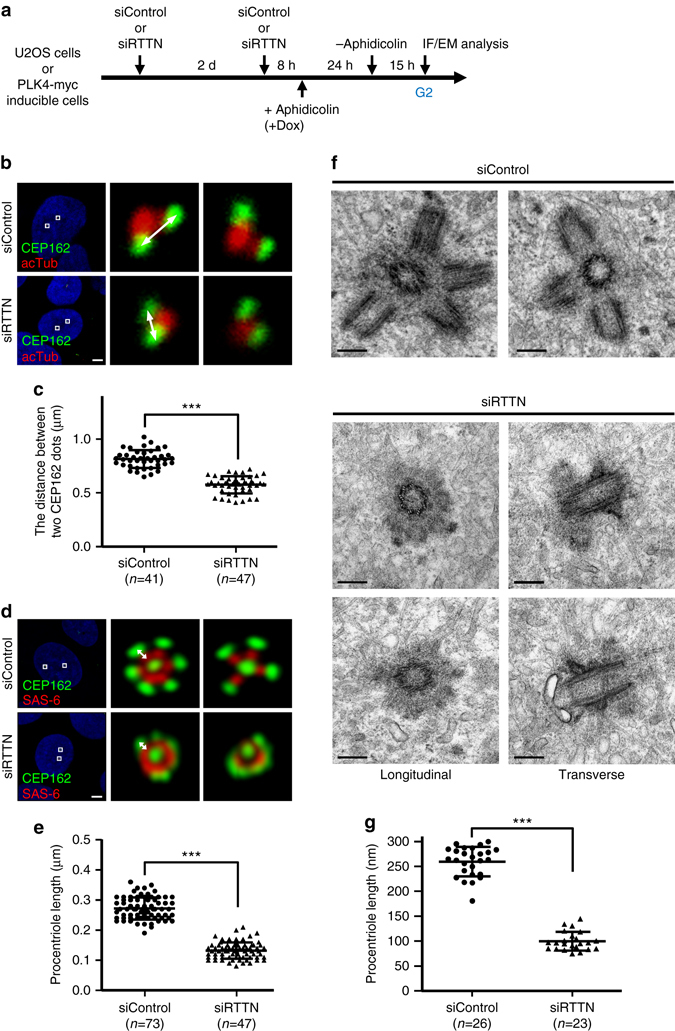
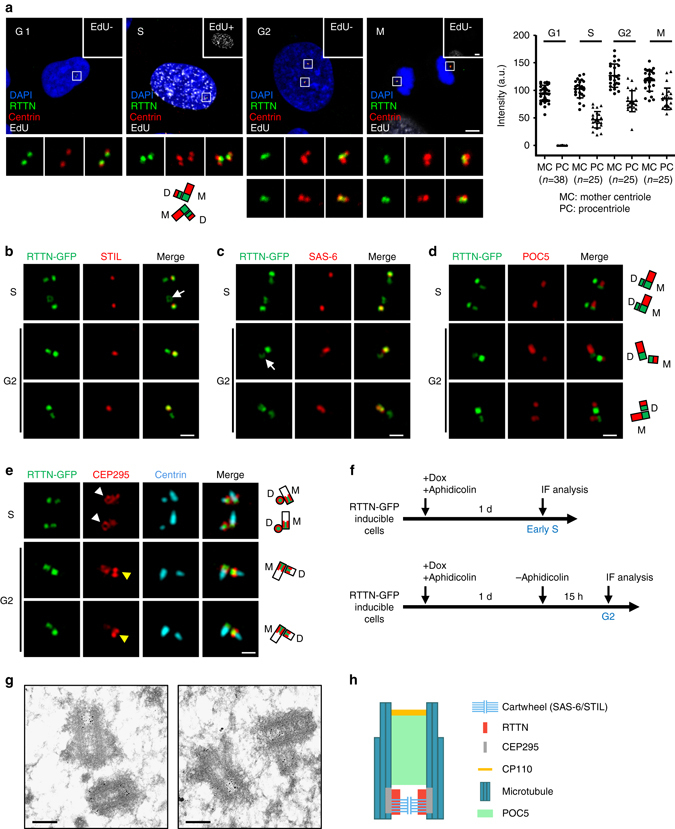
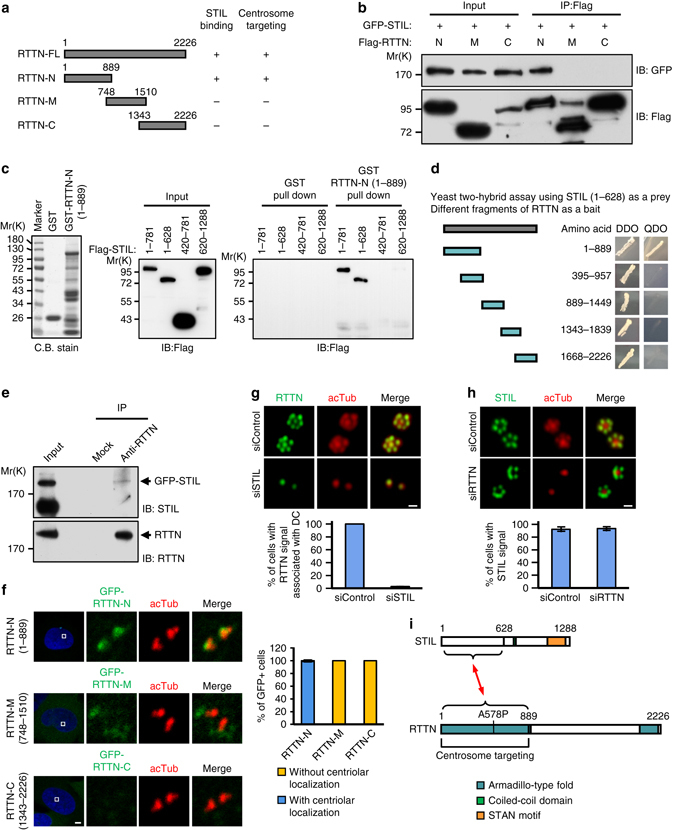
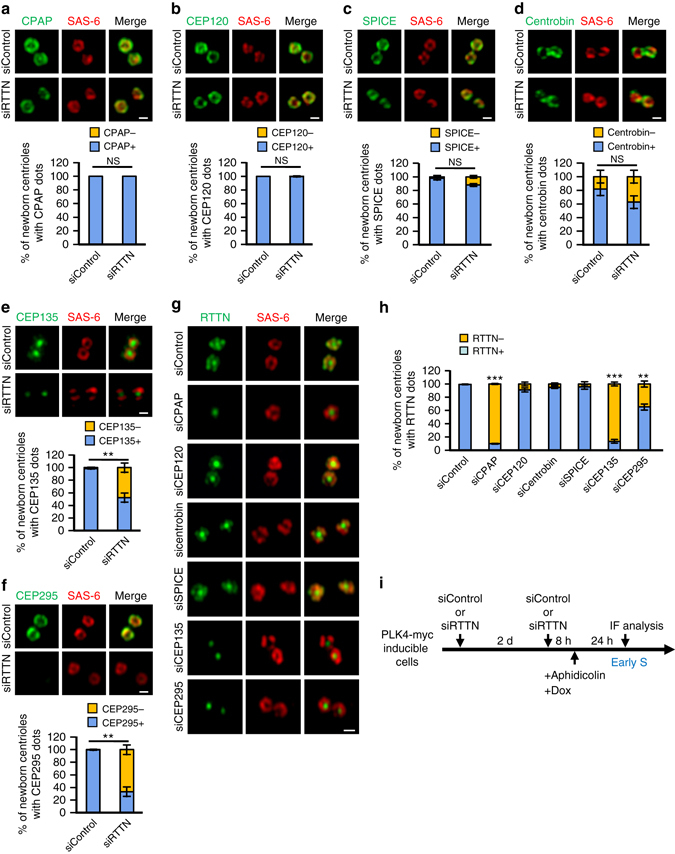
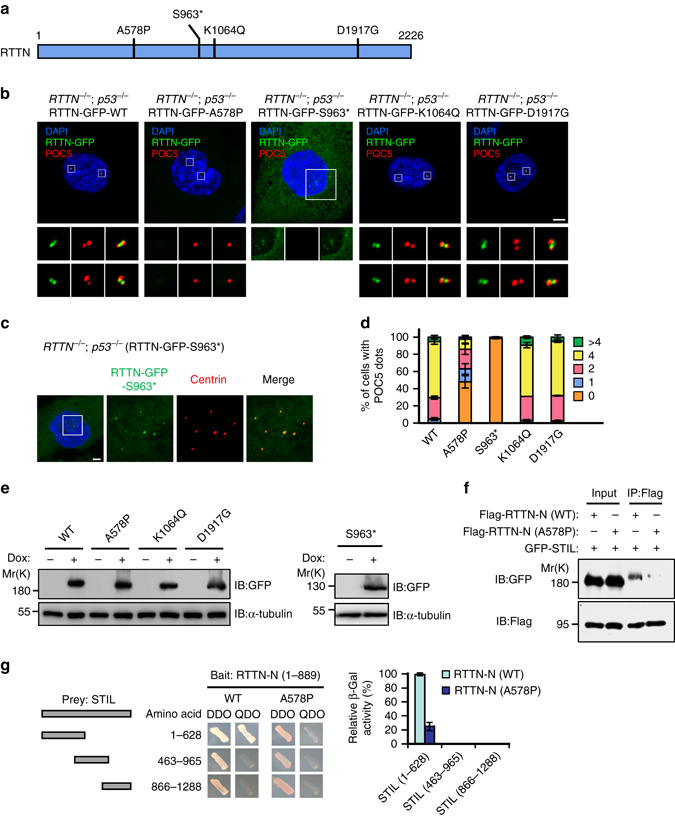
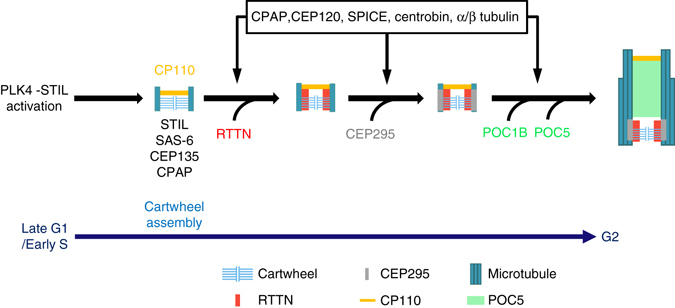
Mutations in many centriolar protein-encoding genes cause primary microcephaly. Using super-resolution and electron microscopy, we find that the human microcephaly protein, RTTN, is recruited to the proximal end of the procentriole at early S phase, and is located at the inner luminal walls of centrioles. Further studies demonstrate that RTTN directly interacts with STIL and acts downstream of STIL-mediated centriole assembly. CRISPR/Cas9-mediated RTTN gene knockout in p53-deficient cells induce amplification of primitive procentriole bodies that lack the distal-half centriolar proteins, POC5 and POC1B. Additional analyses show that RTTN serves as an upstream effector of CEP295, which mediates the loading of POC1B and POC5 to the distal-half centrioles. Interestingly, the naturally occurring microcephaly-associated mutant, RTTN (A578P), shows a low affinity for STIL binding and blocks centriole assembly. These findings reveal that RTTN contributes to building full-length centrioles and illuminate the molecular mechanism through which the RTTN (A578P) mutation causes primary microcephaly.Mutations in many centriolar protein-encoding genes cause primary microcephaly. Here the authors show that human microcephaly protein RTTN directly interacts with STIL and acts downstream of STIL-mediated centriole assembly, contributing to building full-length centrioles.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
