

Inhibition of connexin hemichannels alleviates non-alcoholic steatohepatitis in mice
- PMID: 28811572
- PMCID: PMC5557827
- DOI: 10.1038/s41598-017-08583-w
Inhibition of connexin hemichannels alleviates non-alcoholic steatohepatitis in mice
Abstract
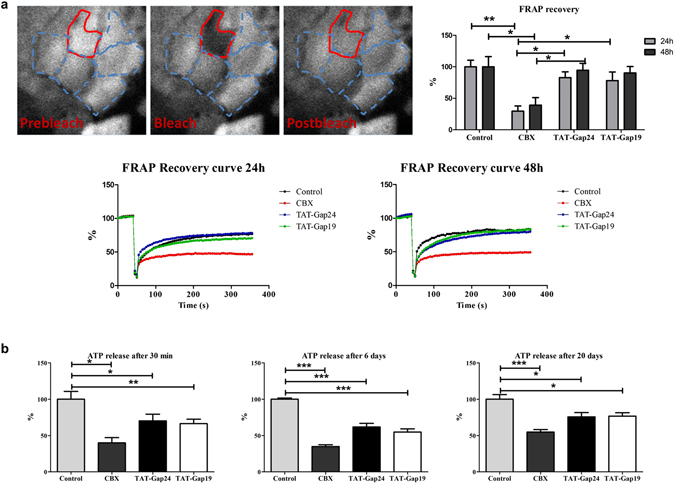
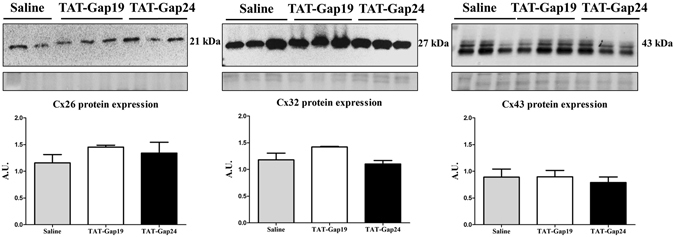
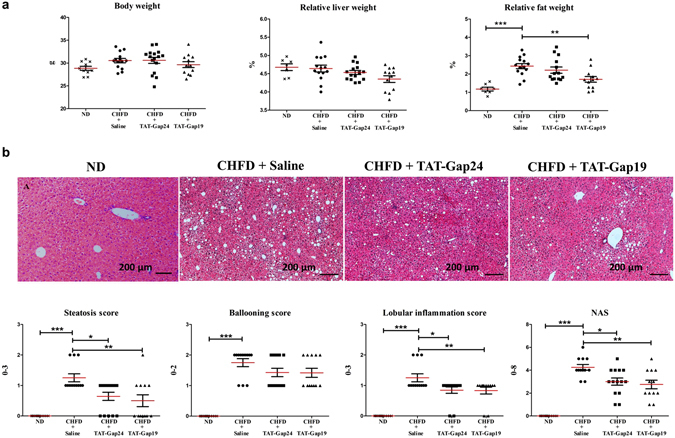
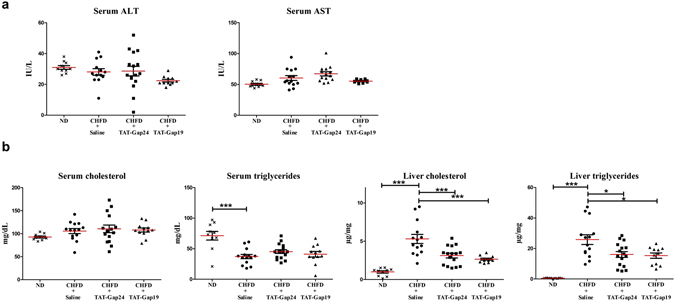
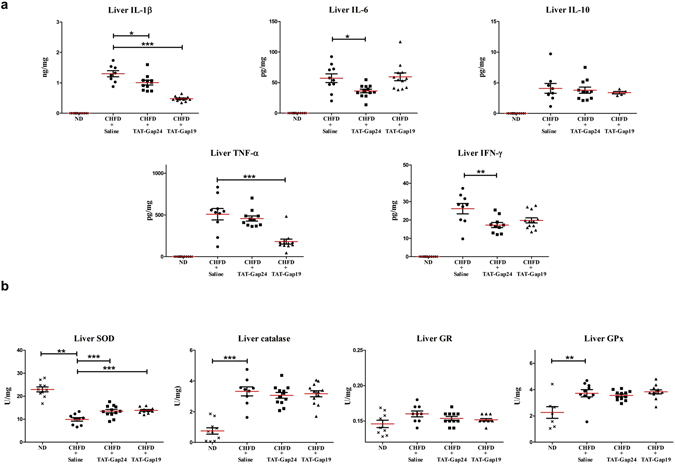
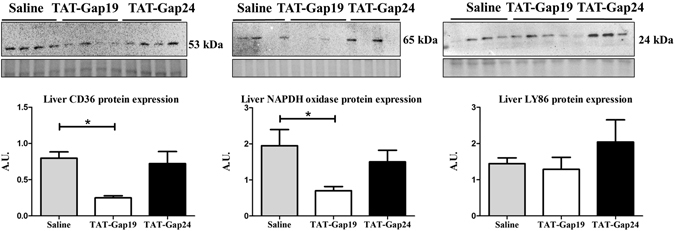
While gap junctions mediate intercellular communication and support liver homeostasis, connexin hemichannels are preferentially opened by pathological stimuli, including inflammation and oxidative stress. The latter are essential features of non-alcoholic steatohepatitis. In this study, it was investigated whether connexin32 and connexin43 hemichannels play a role in non-alcoholic steatohepatitis. Mice were fed a choline-deficient high-fat diet or normal diet for 8 weeks. Thereafter, TAT-Gap24 or TAT-Gap19, specific inhibitors of hemichannels composed of connexin32 and connexin43, respectively, were administered for 2 weeks. Subsequently, histopathological examination was carried out and various indicators of inflammation, liver damage and oxidative stress were tested. In addition, whole transcriptome microarray analysis of liver tissue was performed. Channel specificity of TAT-Gap24 and TAT-Gap19 was examined in vitro by fluorescence recovery after photobleaching analysis and measurement of extracellular release of adenosine triphosphate. TAT-Gap24 and TAT-Gap19 were shown to be hemichannel-specific in cultured primary hepatocytes. Diet-fed animals treated with TAT-Gap24 or TAT-Gap19 displayed decreased amounts of liver lipids and inflammatory markers, and augmented levels of superoxide dismutase, which was supported by the microarray results. These findings show the involvement of connexin32 and connexin43 hemichannels in non-alcoholic steatohepatitis and, simultaneously, suggest a role as potential drug targets in non-alcoholic steatohepatitis.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
