

Substrate-triggered position switching of TatA and TatB during Tat transport in Escherichia coli
- PMID: 28814647
- PMCID: PMC5577447
- DOI: 10.1098/rsob.170091
Substrate-triggered position switching of TatA and TatB during Tat transport in Escherichia coli
Abstract
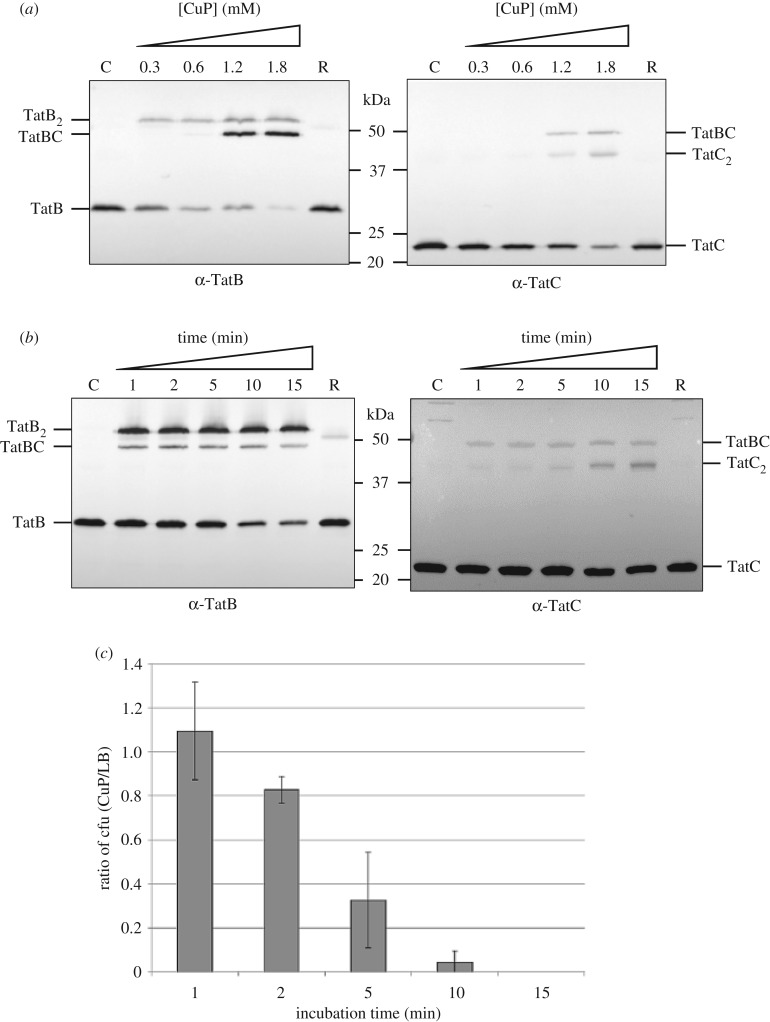
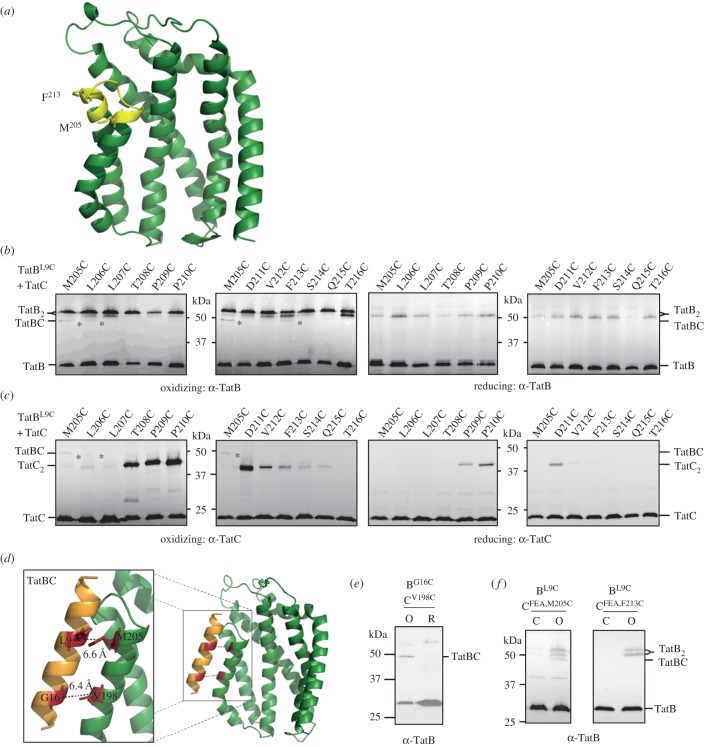
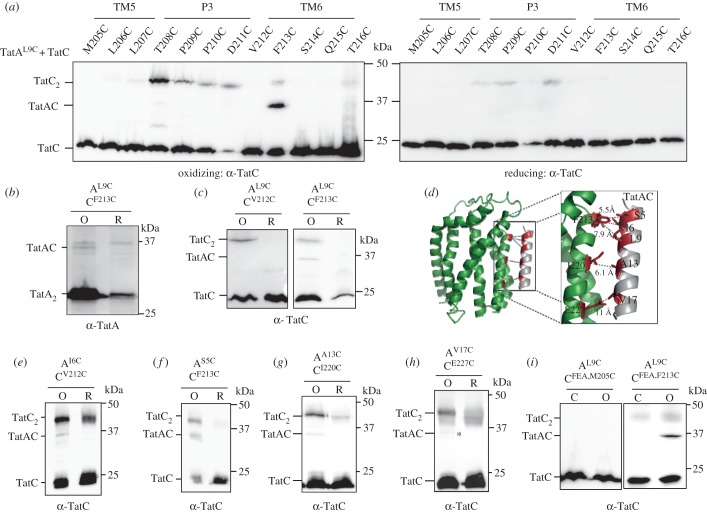
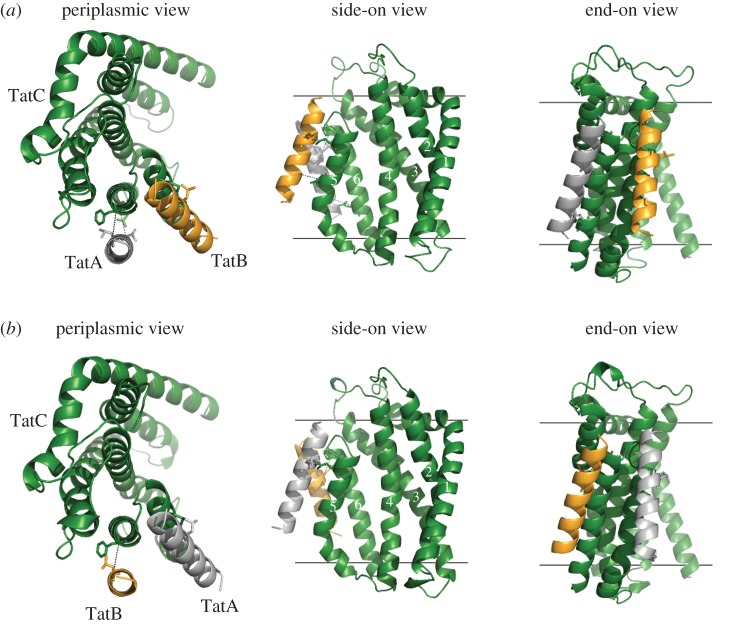
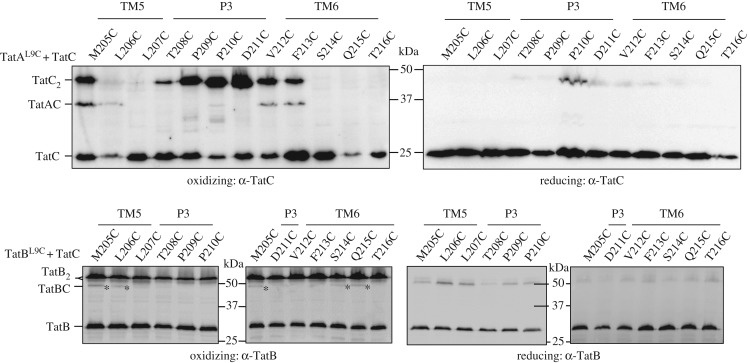
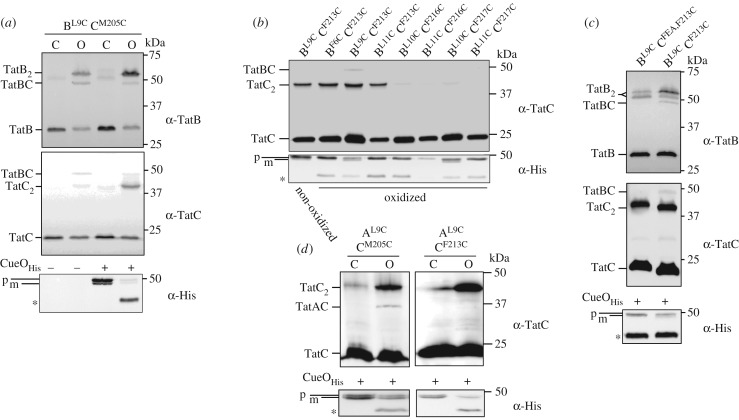
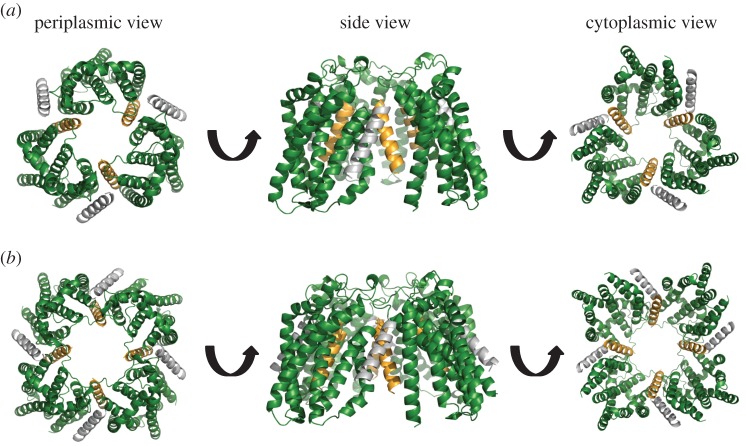
The twin-arginine protein transport (Tat) machinery mediates the translocation of folded proteins across the cytoplasmic membrane of prokaryotes and the thylakoid membrane of plant chloroplasts. The Escherichia coli Tat system comprises TatC and two additional sequence-related proteins, TatA and TatB. The active translocase is assembled on demand, with substrate-binding at a TatABC receptor complex triggering recruitment and assembly of multiple additional copies of TatA; however, the molecular interactions mediating translocase assembly are poorly understood. A 'polar cluster' site on TatC transmembrane (TM) helix 5 was previously identified as binding to TatB. Here, we use disulfide cross-linking and molecular modelling to identify a new binding site on TatC TM helix 6, adjacent to the polar cluster site. We demonstrate that TatA and TatB each have the capacity to bind at both TatC sites, however in vivo this is regulated according to the activation state of the complex. In the resting-state system, TatB binds the polar cluster site, with TatA occupying the TM helix 6 site. However when the system is activated by overproduction of a substrate, TatA and TatB switch binding sites. We propose that this substrate-triggered positional exchange is a key step in the assembly of an active Tat translocase.
Keywords: Tat pathway; protein transport; transport mechanism; twin-arginine signal peptide.
© 2017 The Authors.
Conflict of interest statement
We have no competing interests.
Figures
References
-
- Berks BC. 2015. The twin-arginine protein translocation pathway. Annu. Rev. Biochem. 84, 843–864. (doi:10.1146/annurev-biochem-060614-034251) - DOI - PubMed
-
- Cline K. 2015. Mechanistic aspects of folded protein transport by the twin arginine translocase (Tat) . J. Biol. Chem. 290, 16 530–16 538. (doi:10.1074/jbc.R114.626820) - DOI - PMC - PubMed
-
- Palmer T, Berks BC. 2012. The twin-arginine translocation (Tat) protein export pathway. Nat. Rev. Microbiol. 10, 483–496. (doi:10.1038/nrmicro2814) - DOI - PubMed
-
- Aldridge C, Storm A, Cline K, Dabney-Smith C. 2012. The chloroplast twin arginine transport (Tat) component, Tha4, undergoes conformational changes leading to Tat protein transport. J. Biol. Chem. 287, 34 752–34 763. (doi:10.1074/jbc.M112.385666) - DOI - PMC - PubMed
-
- Koch S, Fritsch MJ, Buchanan G, Palmer T. 2012. The Escherichia coli TatA and TatB proteins have an N-out C-in topology in intact cells. J. Biol. Chem. 287, 14 420–14 431. (doi:10.1074/jbc.M112.354555) - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases