

Gelsolin dysfunction causes photoreceptor loss in induced pluripotent cell and animal retinitis pigmentosa models
- PMID: 28814713
- PMCID: PMC5559447
- DOI: 10.1038/s41467-017-00111-8
Gelsolin dysfunction causes photoreceptor loss in induced pluripotent cell and animal retinitis pigmentosa models
Abstract
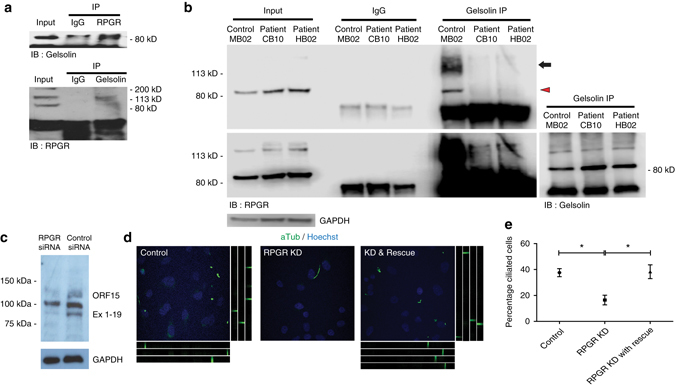
Mutations in the Retinitis Pigmentosa GTPase Regulator (RPGR) cause X-linked RP (XLRP), an untreatable, inherited retinal dystrophy that leads to premature blindness. RPGR localises to the photoreceptor connecting cilium where its function remains unknown. Here we show, using murine and human induced pluripotent stem cell models, that RPGR interacts with and activates the actin-severing protein gelsolin, and that gelsolin regulates actin disassembly in the connecting cilium, thus facilitating rhodopsin transport to photoreceptor outer segments. Disease-causing RPGR mutations perturb this RPGR-gelsolin interaction, compromising gelsolin activation. Both RPGR and Gelsolin knockout mice show abnormalities of actin polymerisation and mislocalisation of rhodopsin in photoreceptors. These findings reveal a clinically-significant role for RPGR in the activation of gelsolin, without which abnormalities in actin polymerisation in the photoreceptor connecting cilia cause rhodopsin mislocalisation and eventual retinal degeneration in XLRP.Mutations in the Retinitis Pigmentosa GTPase Regulator (RPGR) cause retinal dystrophy, but how this arises at a molecular level is unclear. Here, the authors show in induced pluripotent stem cells and mouse knockouts that RPGR mediates actin dynamics in photoreceptors via the actin-severing protein, gelsolin.
Conflict of interest statement
The authors declare no competing financial interests
Figures


References
-
- Besharse J. C. in The Retina: A Model for Cell Biological Studies (eds Adler, R. & Farber, D.) 297–352 (Academic Press, 1986).
Publication types
MeSH terms
Substances
Grants and funding
- G0300336/MRC_/Medical Research Council/United Kingdom
- ERC_/European Research Council/International
- MC_PC_U127584475/MRC_/Medical Research Council/United Kingdom
- MC_UU_12018/26/MRC_/Medical Research Council/United Kingdom
- BB/E012841/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- G0301182/MRC_/Medical Research Council/United Kingdom
- WT_/Wellcome Trust/United Kingdom
- R01 EY022372/EY/NEI NIH HHS/United States
- MC_U127584475/MRC_/Medical Research Council/United Kingdom
- NC/C016106/1/NC3RS_/National Centre for the Replacement, Refinement and Reduction of Animals in Research/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
