

Toward an understanding of the Cdc48/p97 ATPase
- PMID: 28815021
- PMCID: PMC5543420
- DOI: 10.12688/f1000research.11683.1
Toward an understanding of the Cdc48/p97 ATPase
Abstract
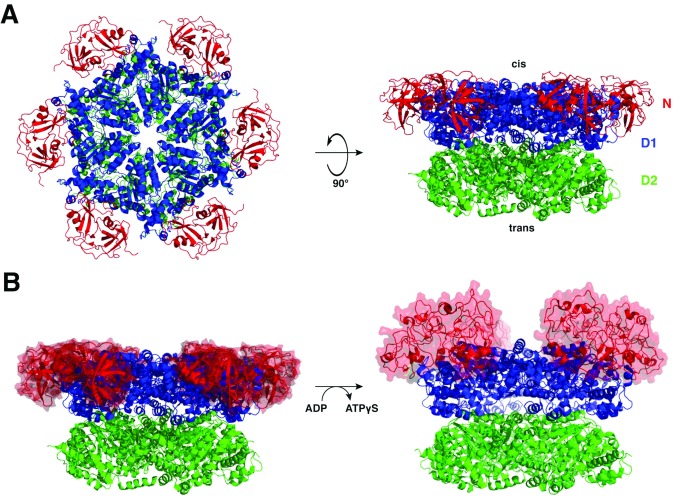
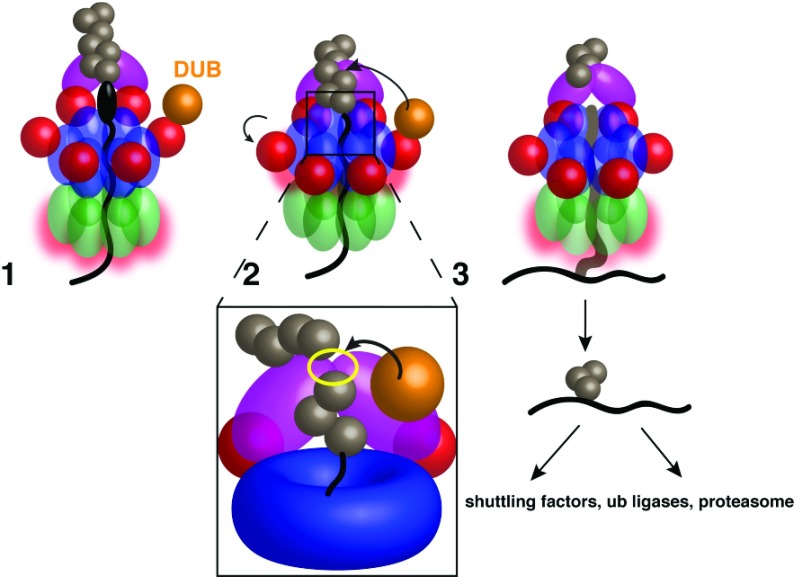
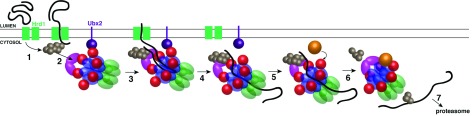
A conserved AAA+ ATPase, called Cdc48 in yeast and p97 or VCP in metazoans, plays an essential role in many cellular processes by segregating polyubiquitinated proteins from complexes or membranes. For example, in endoplasmic reticulum (ER)-associated protein degradation (ERAD), Cdc48/p97 pulls polyubiquitinated, misfolded proteins out of the ER and transfers them to the proteasome. Cdc48/p97 consists of an N-terminal domain and two ATPase domains (D1 and D2). Six Cdc48 monomers form a double-ring structure surrounding a central pore. Cdc48/p97 cooperates with a number of different cofactors, which bind either to the N-terminal domain or to the C-terminal tail. The mechanism of Cdc48/p97 action is poorly understood, despite its critical role in many cellular systems. Recent in vitro experiments using yeast Cdc48 and its heterodimeric cofactor Ufd1/Npl4 (UN) have resulted in novel mechanistic insight. After interaction of the substrate-attached polyubiquitin chain with UN, Cdc48 uses ATP hydrolysis in the D2 domain to move the polypeptide through its central pore, thereby unfolding the substrate. ATP hydrolysis in the D1 domain is involved in substrate release from the Cdc48 complex, which requires the cooperation of the ATPase with a deubiquitinase (DUB). Surprisingly, the DUB does not completely remove all ubiquitin molecules; the remaining oligoubiquitin chain is also translocated through the pore. Cdc48 action bears similarities to the translocation mechanisms employed by bacterial AAA ATPases and the eukaryotic 19S subunit of the proteasome, but differs significantly from that of a related type II ATPase, the NEM-sensitive fusion protein (NSF). Many questions about Cdc48/p97 remain unanswered, including how it handles well-folded substrate proteins, how it passes substrates to the proteasome, and how various cofactors modify substrates and regulate its function.
Keywords: ATPase; Cdc48; p97.
Conflict of interest statement
Competing interests: The authors declare that they have no competing interests.No competing interests were disclosed.No competing interests were disclosed.No competing interests were disclosed.No competing interests were disclosed.No competing interests were disclosed.
Figures

References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous