

Toward a Wiring Diagram Understanding of Appetite Control
- PMID: 28817798
- PMCID: PMC5657399
- DOI: 10.1016/j.neuron.2017.06.014
Toward a Wiring Diagram Understanding of Appetite Control
Abstract
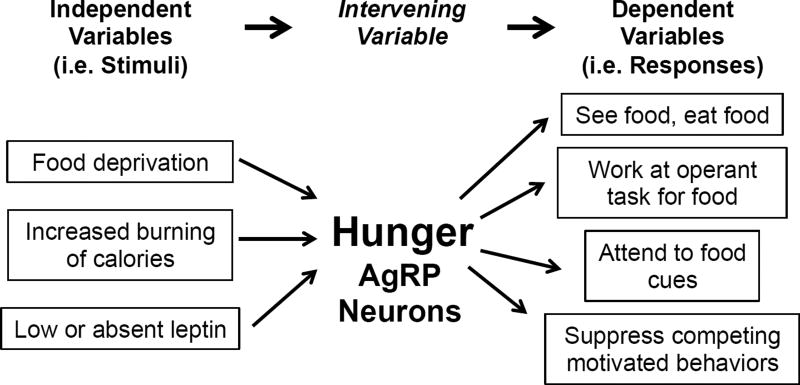
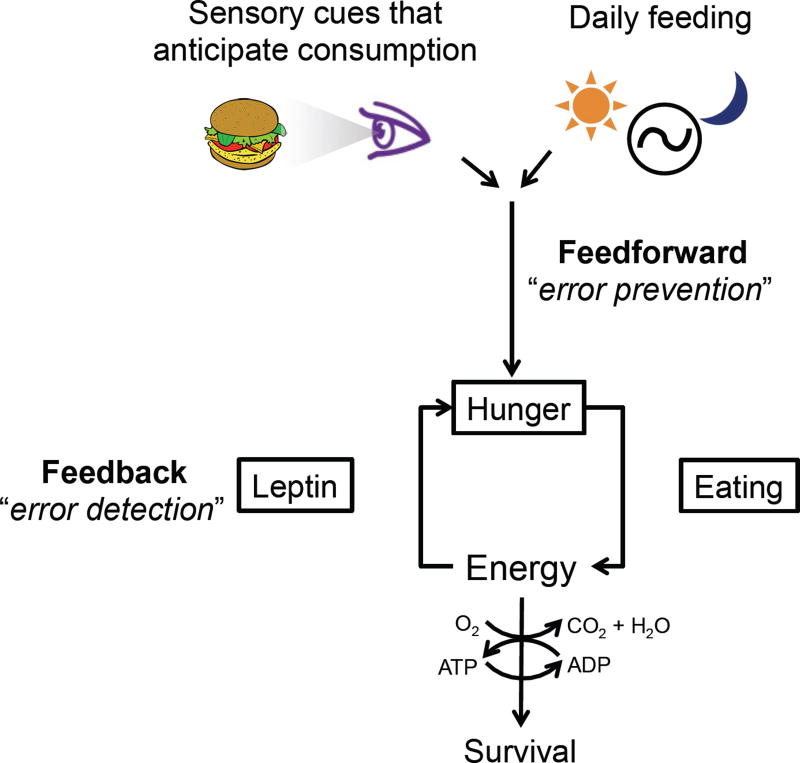
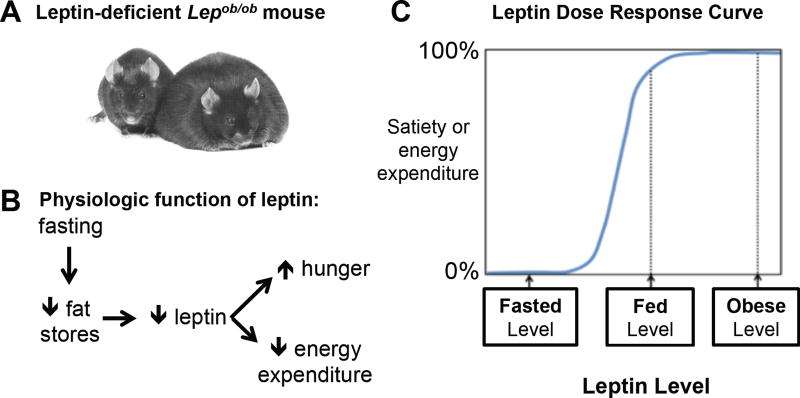
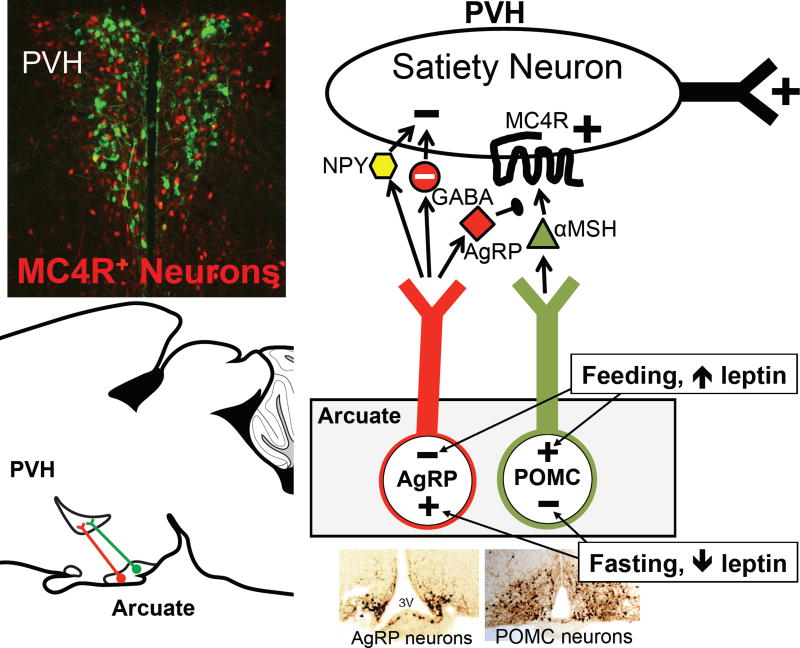
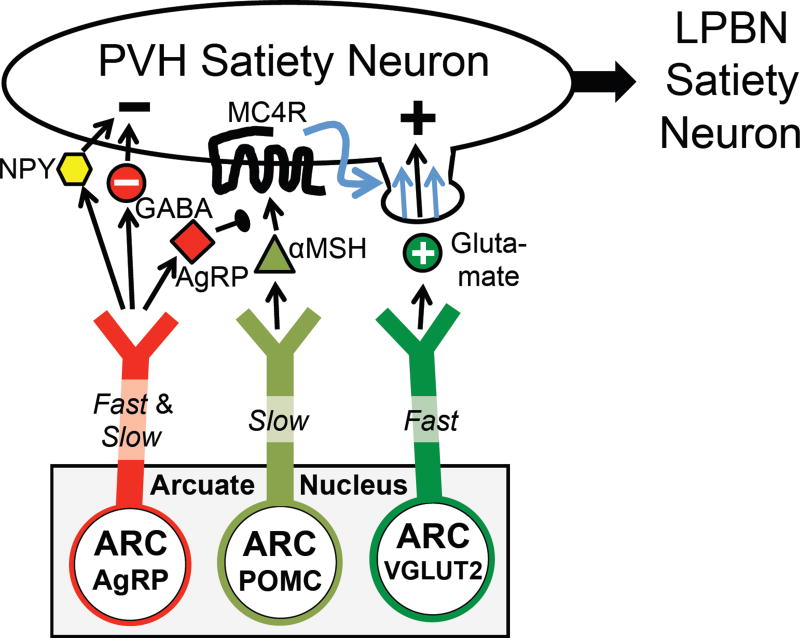
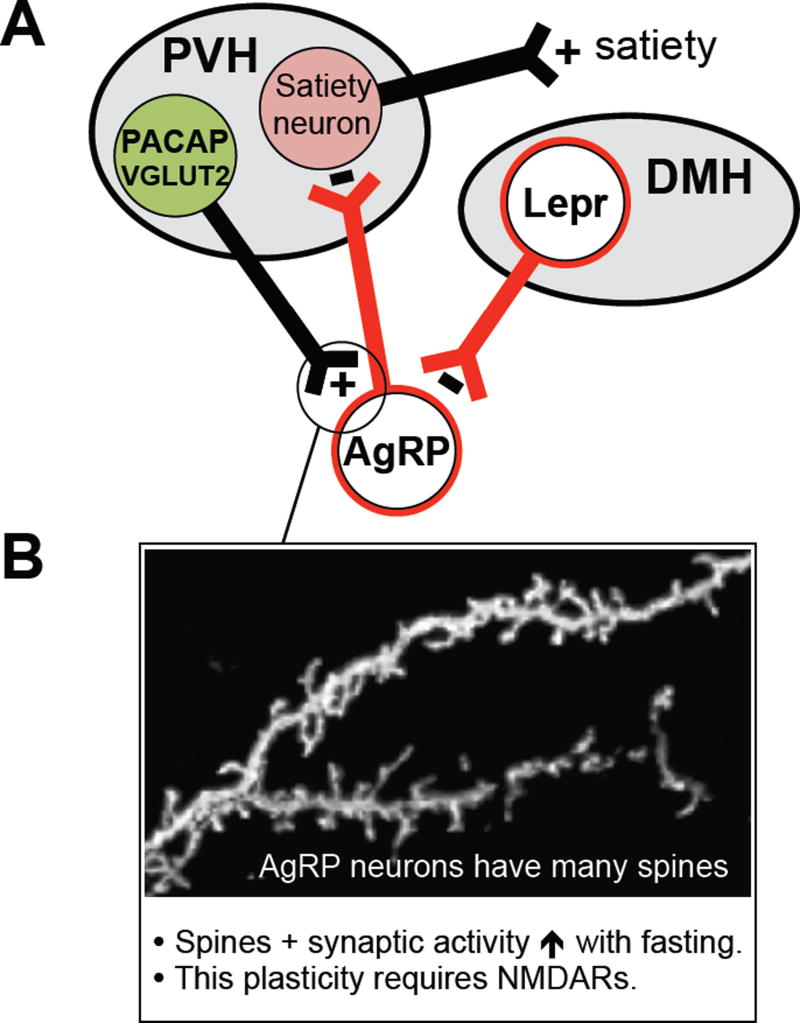
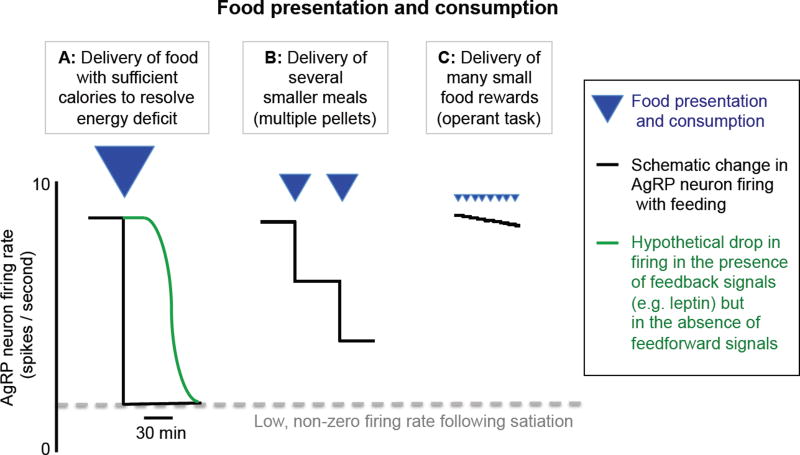
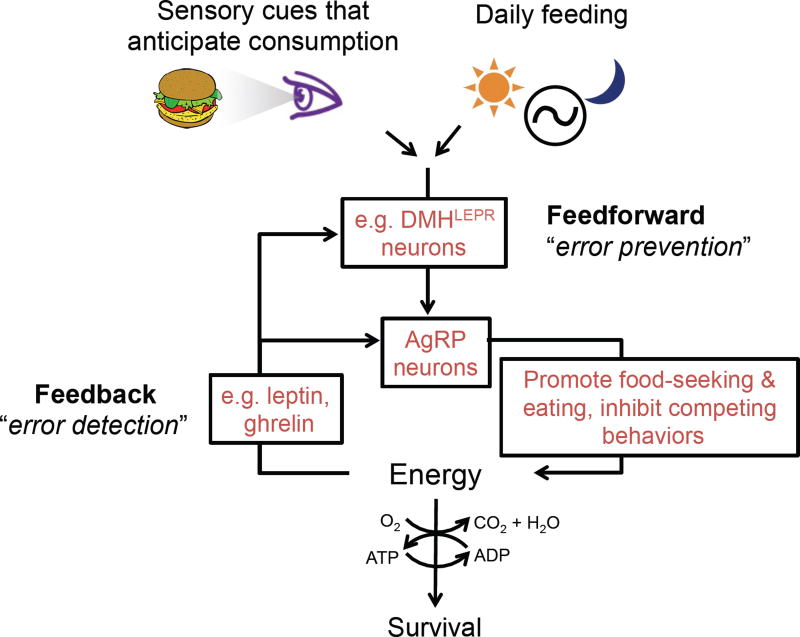
Prior mouse genetic research has set the stage for a deep understanding of appetite regulation. This goal is now being realized through the use of recent technological advances, such as the ability to map connectivity between neurons, manipulate neural activity in real time, and measure neural activity during behavior. Indeed, major progress has been made with regard to meal-related gut control of appetite, arcuate nucleus-based hypothalamic circuits linking energy state to the motivational drive, hunger, and, finally, limbic and cognitive processes that bring about hunger-mediated increases in reward value and perception of food. Unexpected findings are also being made; for example, the rapid regulation of homeostatic neurons by cues that predict future food consumption. The aim of this review is to cover the major underpinnings of appetite regulation, describe recent advances resulting from new technologies, and synthesize these findings into an updated view of appetite regulation.
Keywords: AgRP neurons; POMC neurons; anticipatory regulation; arcuate nucleus; feedforward regulation; food cues; hunger; intervening variable; melanocortin-4 receptor; satiety.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures




References
-
- Ahima RS, Prabakaran D, Mantzoros C, Qu D, Lowell B, Maratos-Flier E, Flier JS. Role of leptin in the neuroendocrine response to fasting. Nature. 1996;382:250–252. - PubMed
-
- Allen LG, Kalra PS, Crowley WR, Kalra SP. Comparison of the effects of neuropeptide Y and adrenergic transmitters on LH release and food intake in male rats. Life Sci. 1985;37:617–623. - PubMed
-
- Aotani D, Ebihara K, Sawamoto N, Kusakabe T, Aizawa-Abe M, Kataoka S, Sakai T, Iogawa H, Ebihara C, Fujikura J, et al. Functional magnetic resonance imaging analysis of food-related brain activity in patients with lipodystrophy undergoing leptin replacement therapy. J Clin Endocrinol Metab. 2012;97:3663–3671. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
