

Sexual dimorphism of the pulmonary transcriptome in neonatal hyperoxic lung injury: identification of angiogenesis as a key pathway
- PMID: 28818871
- PMCID: PMC5814706
- DOI: 10.1152/ajplung.00230.2017
Sexual dimorphism of the pulmonary transcriptome in neonatal hyperoxic lung injury: identification of angiogenesis as a key pathway
Abstract
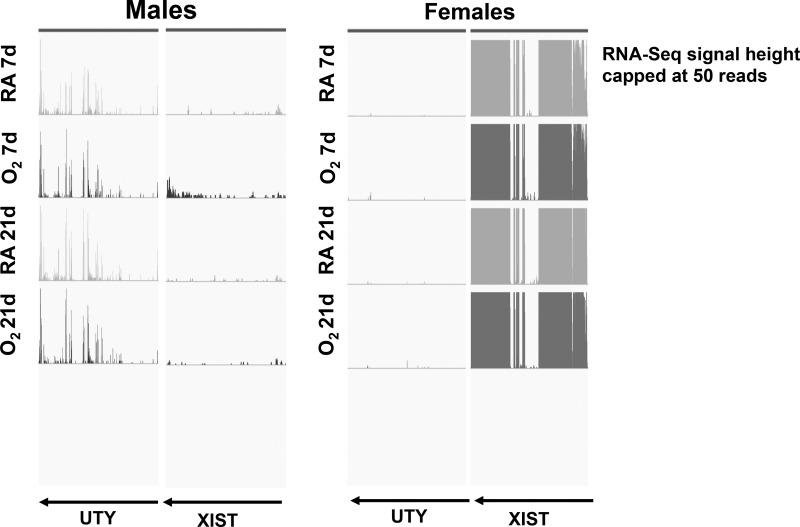
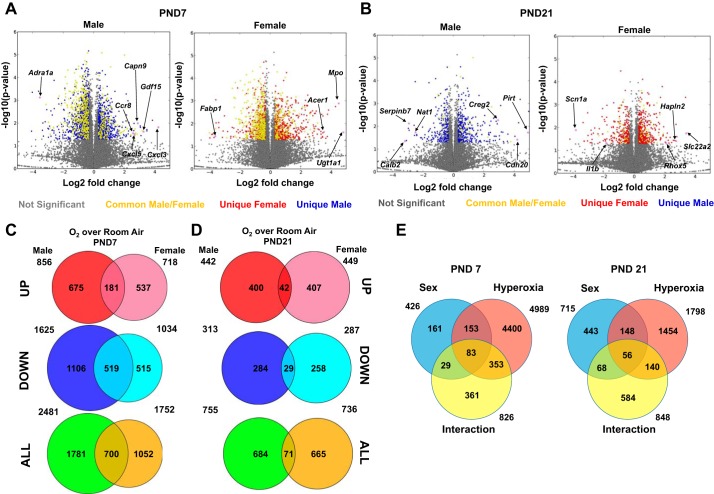
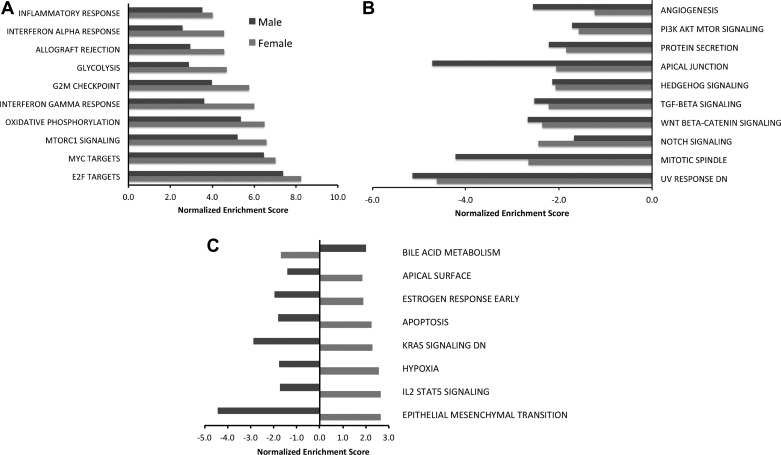
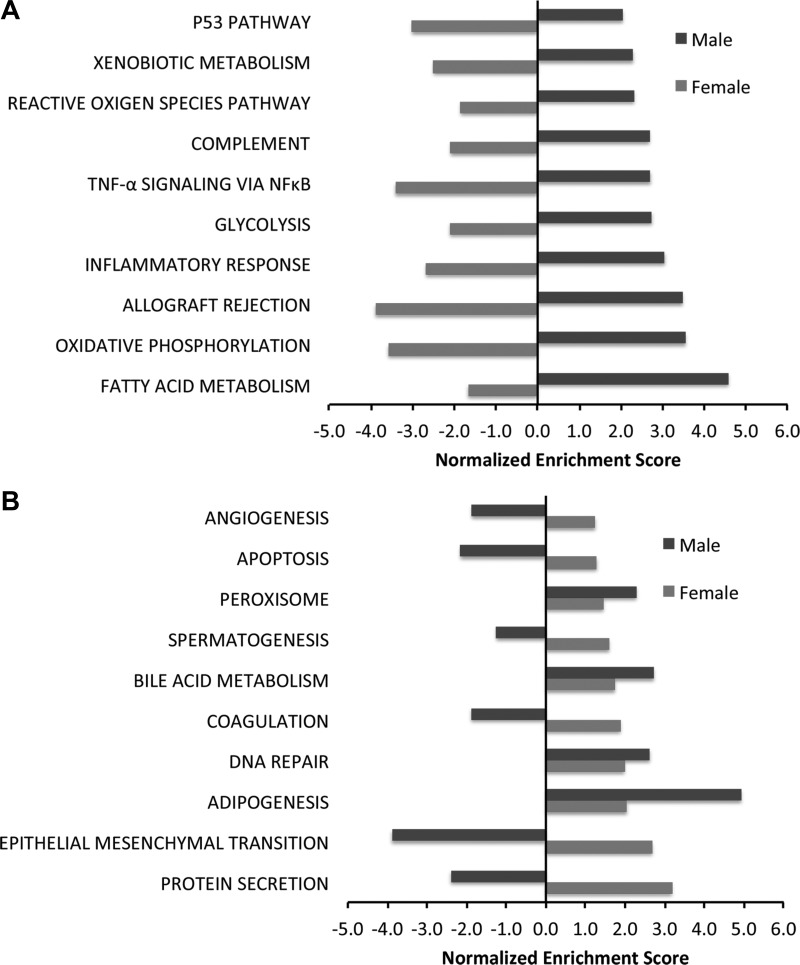
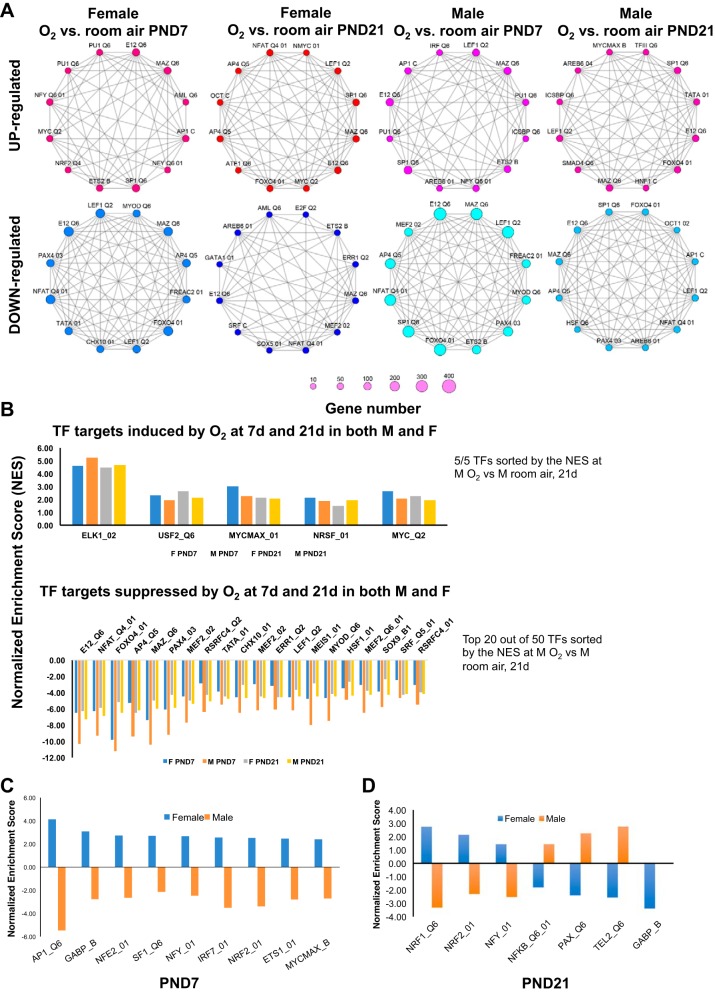
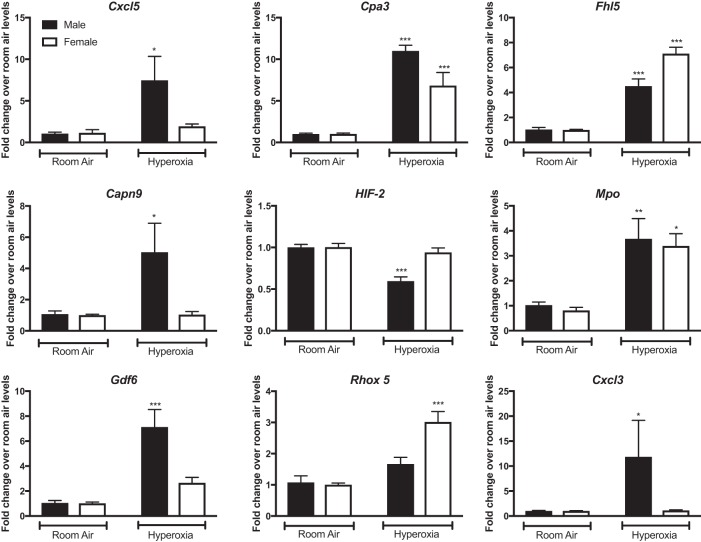
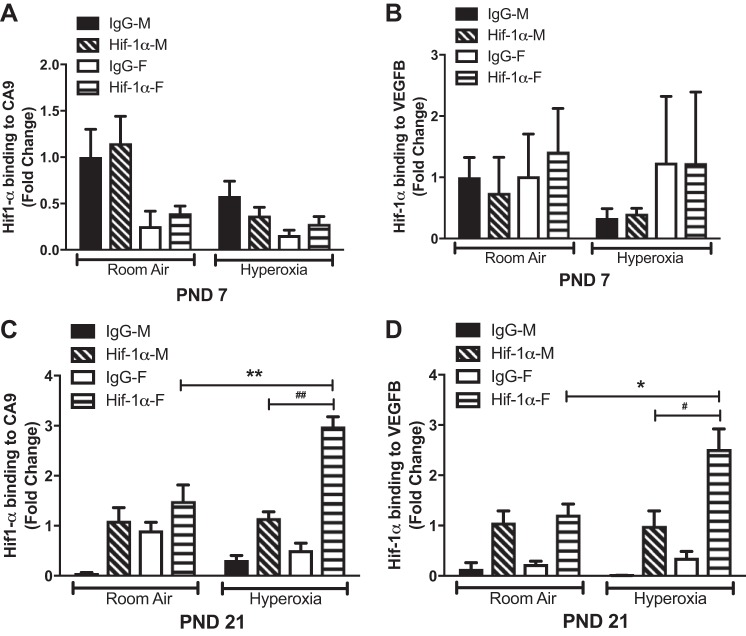
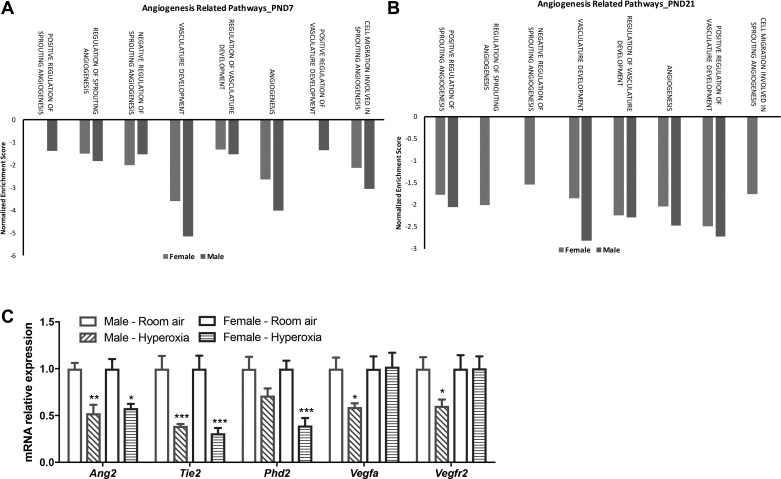
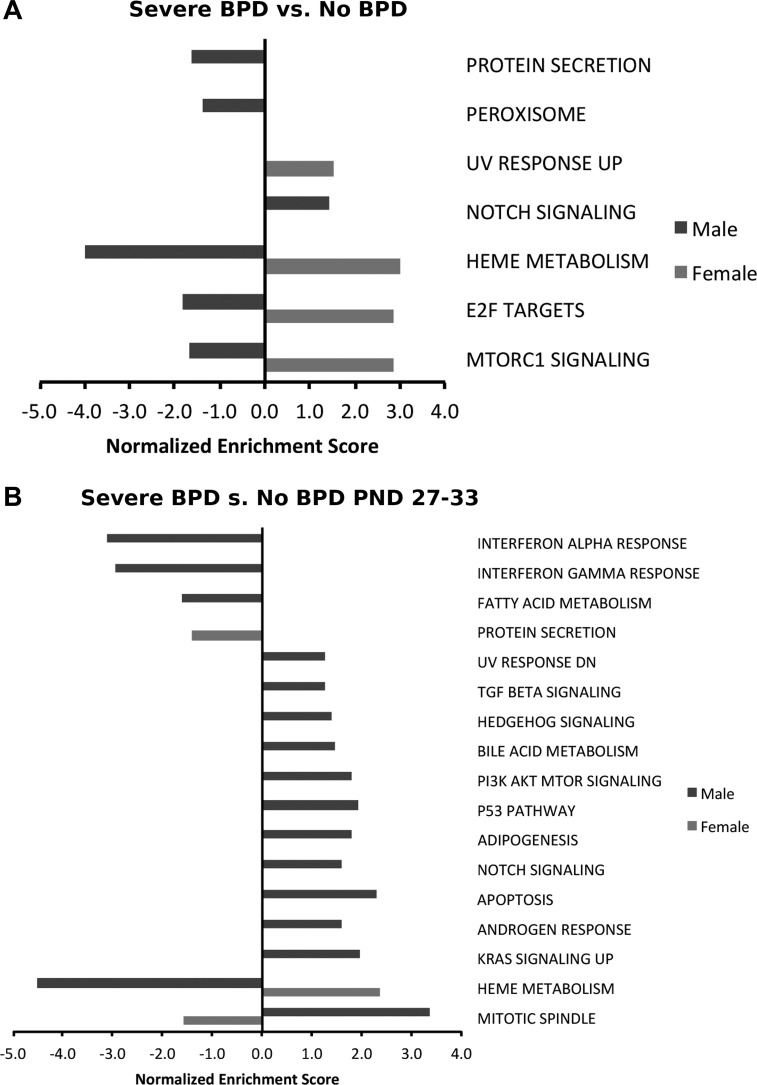
Bronchopulmonary dysplasia (BPD) is characterized by impaired alveolar secondary septation and vascular growth. Exposure to high concentrations of oxygen (hyperoxia) contributes to the development of BPD. The male sex is considered an independent risk factor for the development of BPD. The reasons underlying sexually dimorphic outcomes in premature neonates are not known. We hypothesized that sex-specific modulation of biological processes in the lung under hyperoxic conditions contributes to sex-based differences. Neonatal male and female mice (C57BL/6) were exposed to hyperoxia [95% [Formula: see text], postnatal day (PND) 1-5: saccular stage of lung development] and euthanized on PND 7 or 21. Pulmonary gene expression was studied using RNA-Seq on the Illumina HiSeq 2500 platform. Analysis of the pulmonary transcriptome revealed differential sex-specific modulation of crucial pathways such as angiogenesis, response to hypoxia, inflammatory response, and p53 pathway. Candidate genes from these pathways were validated at the mRNA level by qPCR. Analysis also revealed sex-specific differences in the modulation of crucial transcription factors. Focusing on the differential modulation of the angiogenesis pathway, we also showed sex-specific differential activation of Hif-1α-regulated genes using ChIP-qPCR and differences in expression of crucial genes (Vegf, VegfR2, and Phd2) modulating angiogenesis. We demonstrate the translational relevance of our findings by showing that our murine sex-specific differences in gene expression correlate with those from a preexisting human BPD data set. In conclusion, we provide novel molecular insights into differential sex-specific modulation of the pulmonary transcriptome in neonatal hyperoxic lung injury and highlight angiogenesis as one of the crucial differentially modulated pathways.
Keywords: RNA-Seq; bronchopulmonary dysplasia; hyperoxia; sex.
Copyright © 2017 the American Physiological Society.
Figures
References
-
- Asikainen TM, Schneider BK, Waleh NS, Clyman RI, Ho W-B, Flippin LA, Günzler V, White CW. Activation of hypoxia-inducible factors in hyperoxia through prolyl 4-hydroxylase blockade in cells and explants of primate lung. Proc Natl Acad Sci USA 102: 10212–10217, 2005. doi: 10.1073/pnas.0504520102. - DOI - PMC - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous