

Mutations in the promoter of the telomerase gene TERT contribute to tumorigenesis by a two-step mechanism
- PMID: 28818973
- PMCID: PMC5942222
- DOI: 10.1126/science.aao0535
Mutations in the promoter of the telomerase gene TERT contribute to tumorigenesis by a two-step mechanism
Abstract
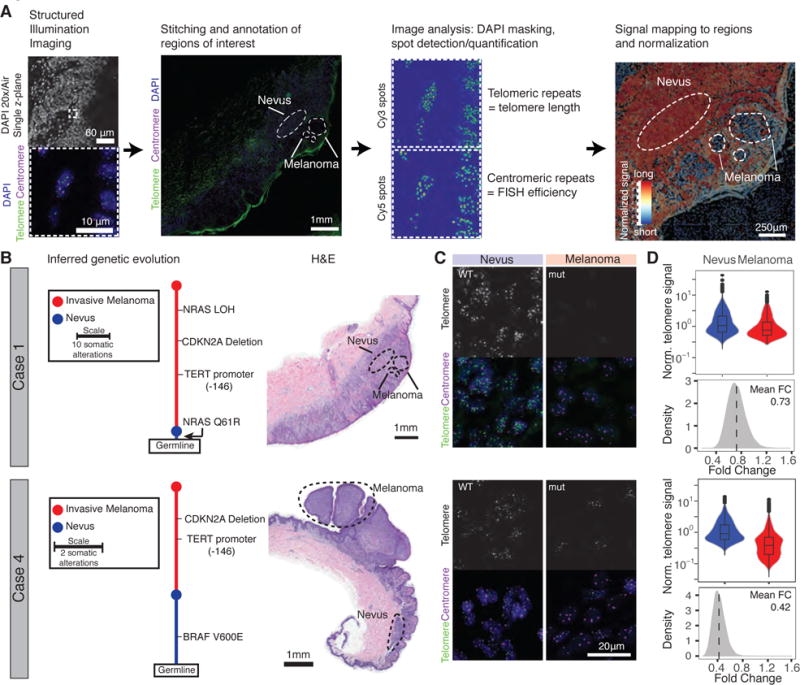
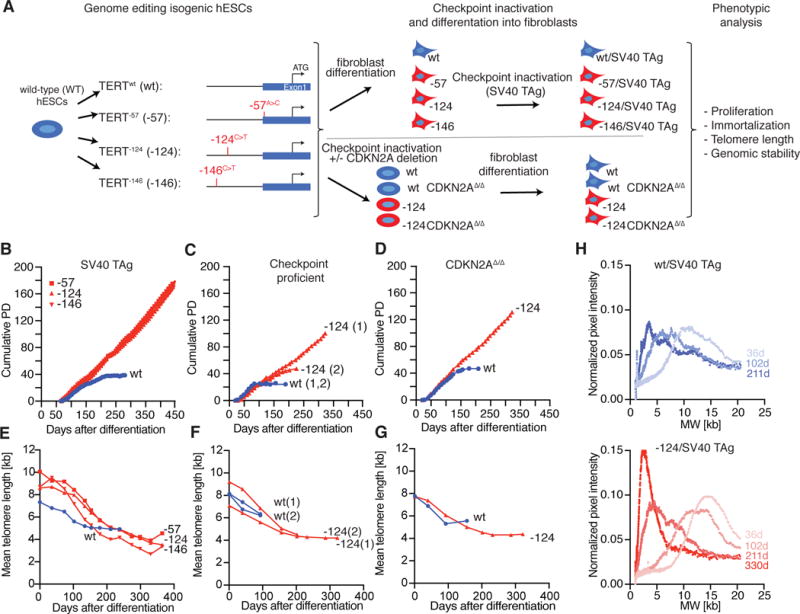
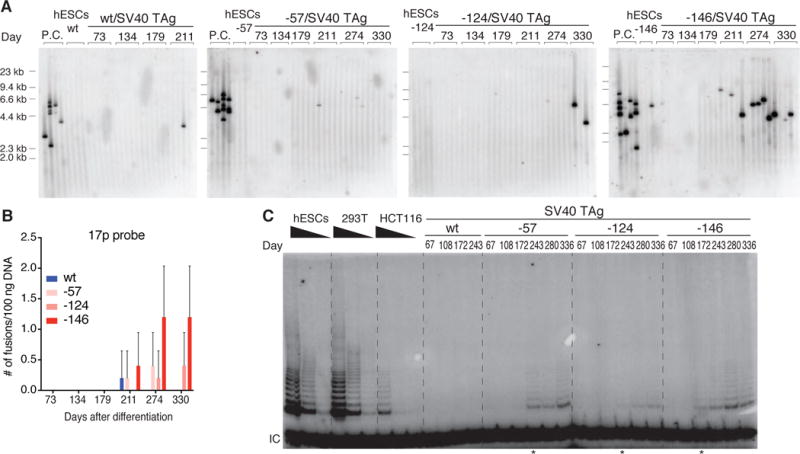
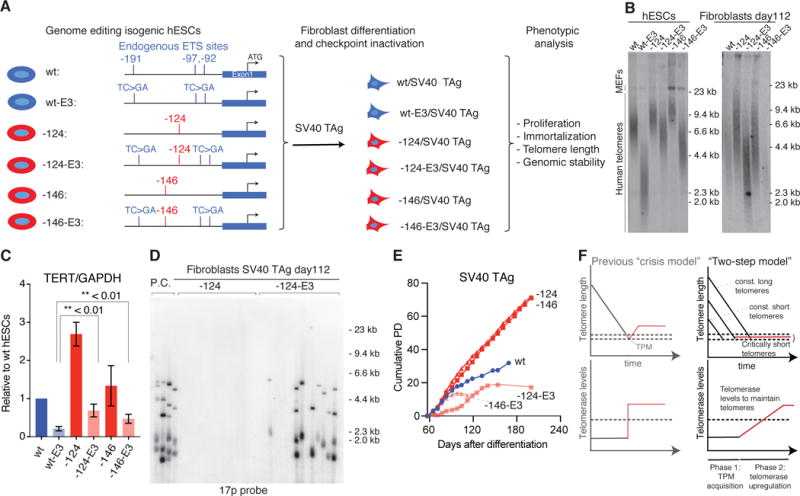
TERT promoter mutations (TPMs) are the most common noncoding mutations in cancer. The timing and consequences of TPMs have not been fully established. Here, we show that TPMs acquired at the transition from benign nevus to malignant melanoma do not support telomere maintenance. In vitro experiments revealed that TPMs do not prevent telomere attrition, resulting in cells with critically short and unprotected telomeres. Immortalization by TPMs requires a gradual up-regulation of telomerase, coinciding with telomere fusions. These data suggest that TPMs contribute to tumorigenesis by promoting immortalization and genomic instability in two phases. In an initial phase, TPMs do not prevent bulk telomere shortening but extend cellular life span by healing the shortest telomeres. In the second phase, the critically short telomeres lead to genome instability and telomerase is further up-regulated to sustain cell proliferation.
Copyright © 2017 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
Comment in
-
New insights into melanoma development.Science. 2017 Sep 29;357(6358):1358-1359. doi: 10.1126/science.aao6963. Epub 2017 Sep 28. Science. 2017. PMID: 28963245 No abstract available.
References
-
- Greider CW, Blackburn EH. Identification of a specific telomere terminal transferase activity in Tetrahymena extracts. Cell. 1985;43:405–413. - PubMed
-
- Greider CW, Blackburn EH. The telomere terminal transferase of Tetrahymena is a ribonucleoprotein enzyme with two kinds of primer specificity. Cell. 1987;51:887–898. - PubMed
-
- Horn S, et al. TERT promoter mutations in familial and sporadic melanoma. Science. 2013;339:959–961. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
