

Landscape of submitochondrial protein distribution
- PMID: 28819139
- PMCID: PMC5561175
- DOI: 10.1038/s41467-017-00359-0
Landscape of submitochondrial protein distribution
Abstract
The mitochondrial proteome comprises ~1000 (yeast)-1500 (human) different proteins, which are distributed into four different subcompartments. The sublocalization of these proteins within the organelle in most cases remains poorly defined. Here we describe an integrated approach combining stable isotope labeling, various protein enrichment and extraction strategies and quantitative mass spectrometry to produce a quantitative map of submitochondrial protein distribution in S. cerevisiae. This quantitative landscape enables a proteome-wide classification of 986 proteins into soluble, peripheral, and integral mitochondrial membrane proteins, and the assignment of 818 proteins into the four subcompartments: outer membrane, inner membrane, intermembrane space, or matrix. We also identified 206 proteins that were not previously annotated as localized to mitochondria. Furthermore, the protease Prd1, misannotated as intermembrane space protein, could be re-assigned and characterized as a presequence peptide degrading enzyme in the matrix.Protein localization plays an important role in the regulation of cellular physiology. Here the authors use an integrated proteomics approach to localize proteins to the mitochondria and provide a detailed map of their specific localization within the organelle.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
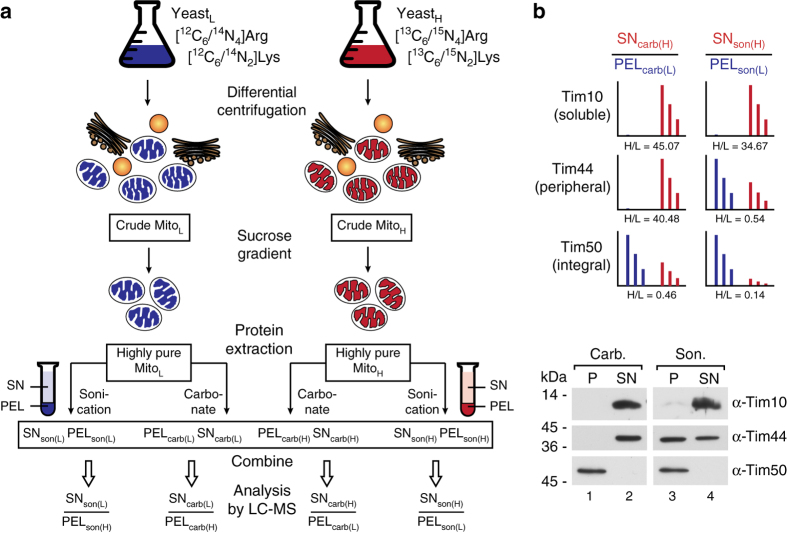
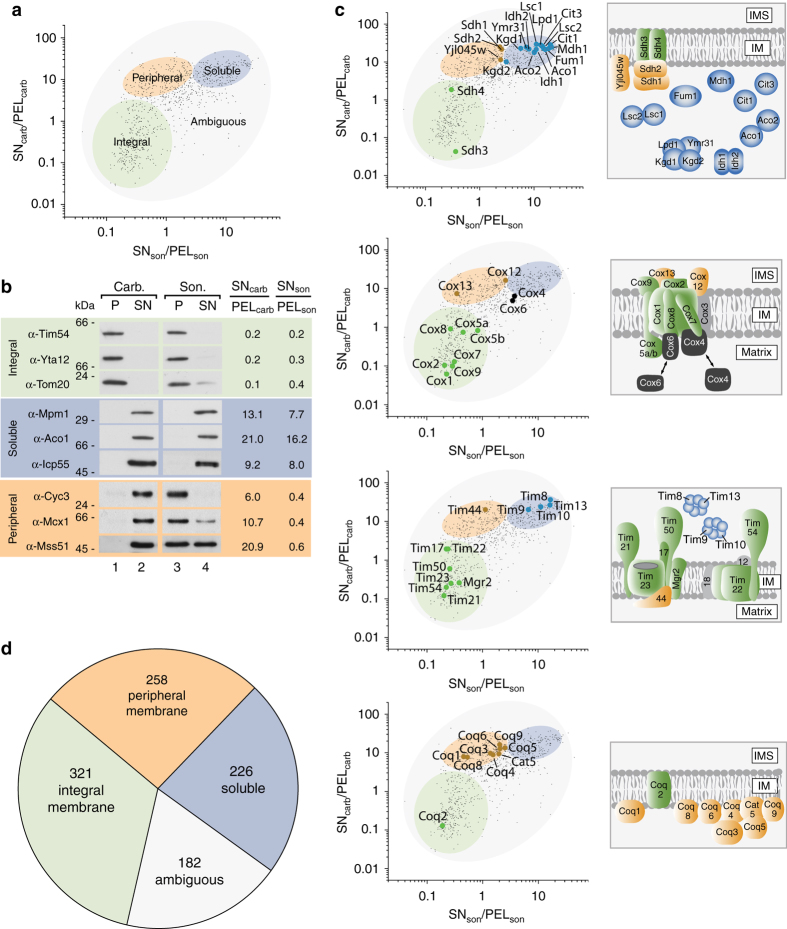
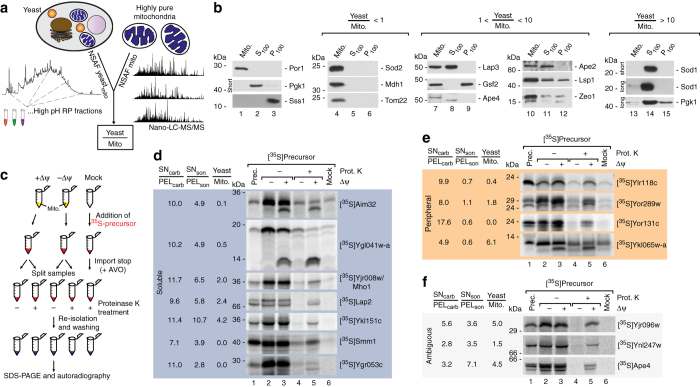
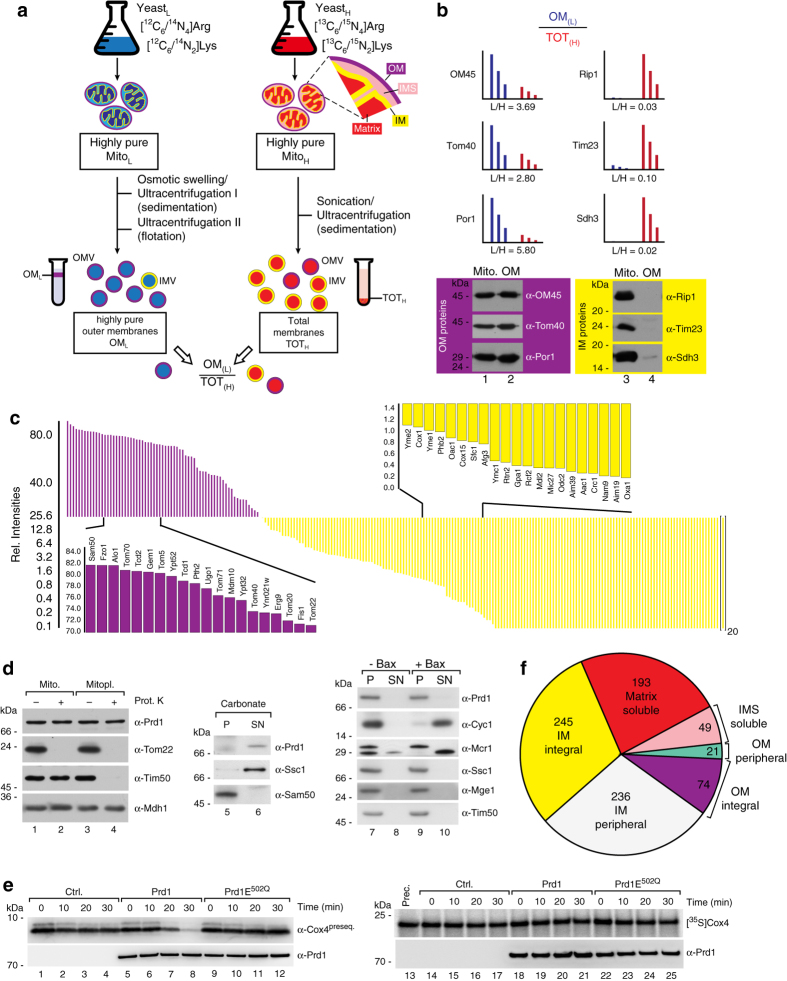
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
