

Dynamic imaging of mitochondrial membrane proteins in specific sub-organelle membrane locations
- PMID: 28819924
- PMCID: PMC5578930
- DOI: 10.1007/s12551-017-0287-1
Dynamic imaging of mitochondrial membrane proteins in specific sub-organelle membrane locations
Abstract
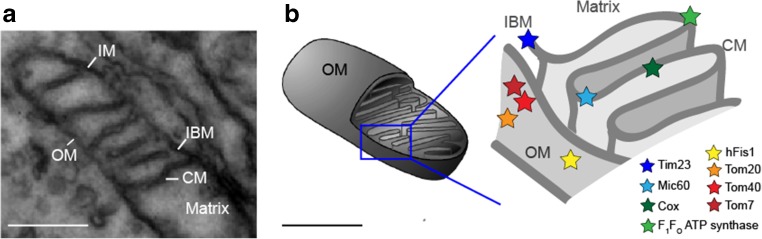
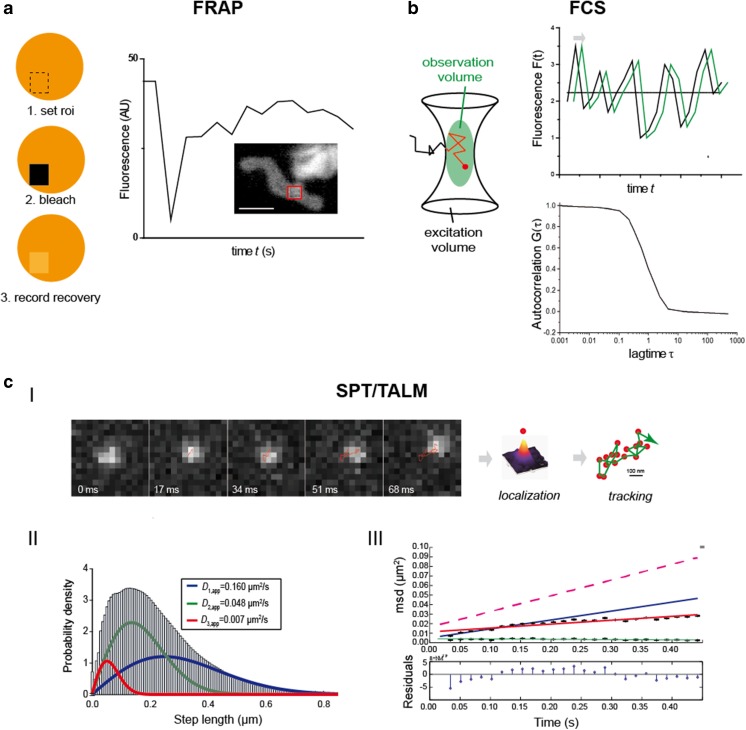
Mitochondria are cellular organelles with multifaceted tasks and thus composed of different sub-compartments. The inner mitochondrial membrane especially has a complex nano-architecture with cristae protruding into the matrix. Related to their function, the localization of mitochondrial membrane proteins is more or less restricted to specific sub-compartments. In contrast, it can be assumed that membrane proteins per se diffuse unimpeded through continuous membranes. Fluorescence recovery after photobleaching is a versatile technology used in mobility analyses to determine the mobile fraction of proteins, but it cannot provide data on subpopulations or on confined diffusion behavior. Fluorescence correlation spectroscopy is used to analyze single molecule diffusion, but no trajectory maps are obtained. Single particle tracking (SPT) technologies in live cells, such as tracking and localization microscopy (TALM), do provide nanotopic localization and mobility maps of mitochondrial proteins in situ. Molecules can be localized with a precision of between 10 and 20 nm, and single trajectories can be recorded and analyzed; this is sufficient to reveal significant differences in the spatio-temporal behavior of diverse mitochondrial proteins. Here, we compare diffusion coefficients obtained by these different technologies and discuss trajectory maps of diverse mitochondrial membrane proteins obtained by SPT/TALM. We show that membrane proteins in the outer membrane generally display unhindered diffusion, while the mobility of inner membrane proteins is restricted by the inner membrane architecture, resulting in significantly lower diffusion coefficients. Moreover, tracking analysis could discern proteins in the inner boundary membrane from proteins preferentially diffusing in cristae membranes, two sub-compartments of the inner mitochondrial membrane. Thus, by evaluating trajectory maps it is possible to assign proteins to different sub-compartments of the same membrane.
Keywords: Cox; F1FO ATP synthase; FCS; FRAP; Membrane sub-compartments; Mitochondrial membrane proteins; Mitofilin/Mic60; Mobility analyses; SPT technologies; Single molecule localization and diffusion; TIM; TOM; hFis.
Conflict of interest statement
Conflict of interest
Timo Appelhans declares that he has no conflicts of interest. Karin B. Busch declares that she has no conflicts of interest.
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Figures

References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
