

TGF-β-induced intracellular PAI-1 is responsible for retaining hematopoietic stem cells in the niche
- PMID: 28821477
- PMCID: PMC5701521
- DOI: 10.1182/blood-2017-02-767384
TGF-β-induced intracellular PAI-1 is responsible for retaining hematopoietic stem cells in the niche
Abstract
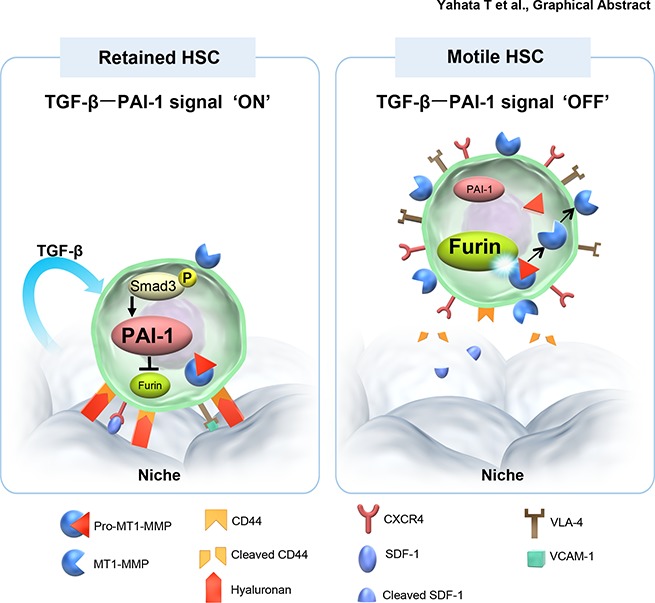
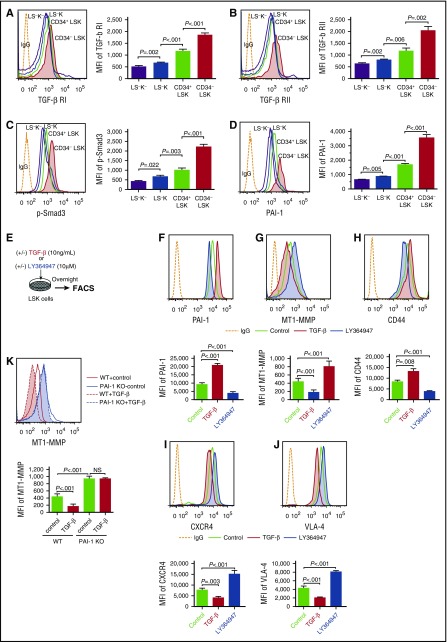
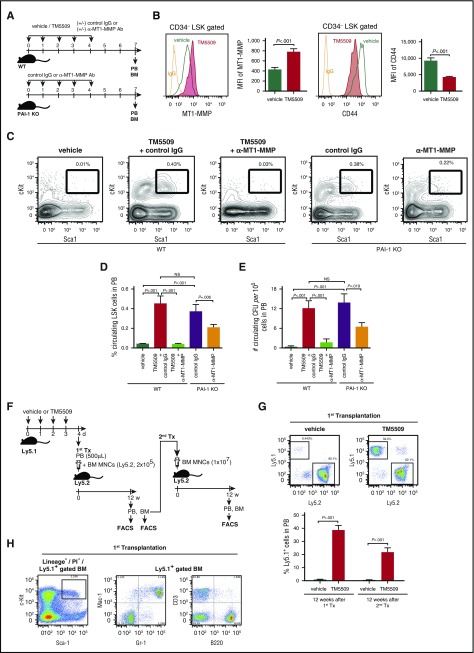
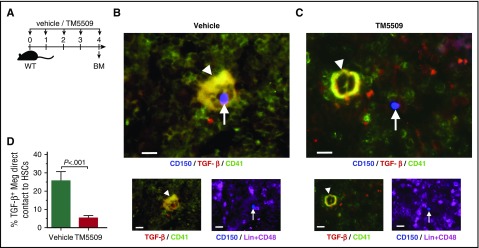
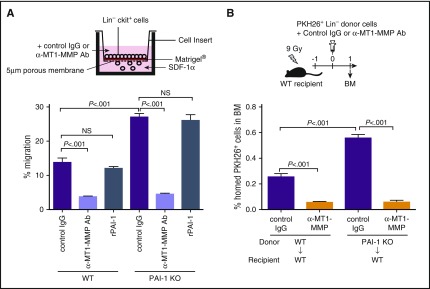
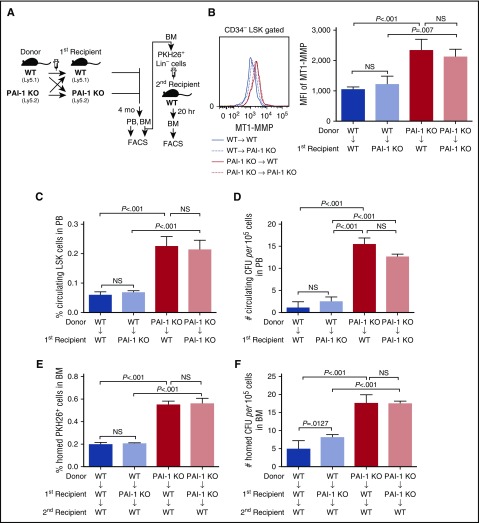
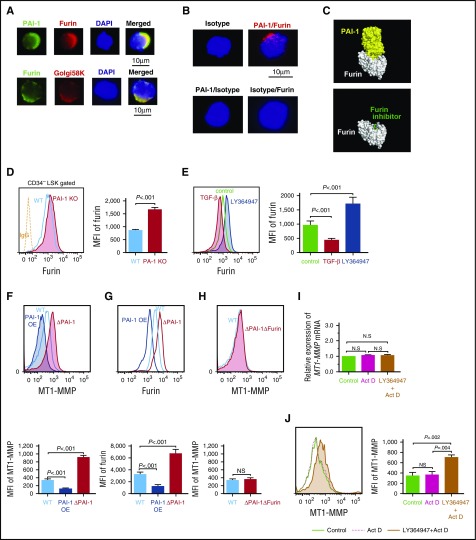
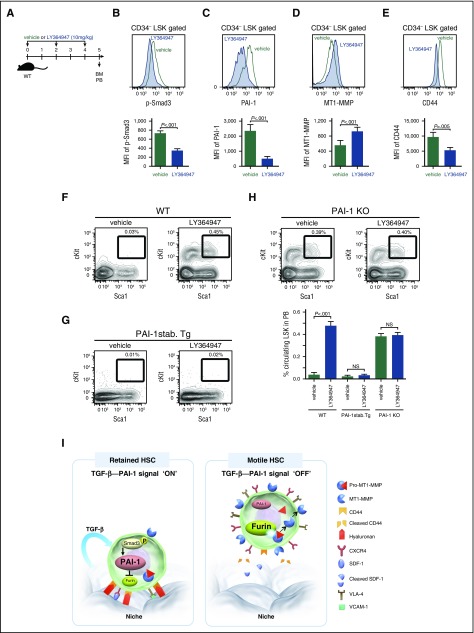
Hematopoietic stem and progenitor cells (HSPCs) reside in the supportive stromal niche in bone marrow (BM); when needed, however, they are rapidly mobilized into the circulation, suggesting that HSPCs are intrinsically highly motile but usually stay in the niche. We questioned what determines the motility of HSPCs. Here, we show that transforming growth factor (TGF)-β-induced intracellular plasminogen activator inhibitor (PAI)-1 activation is responsible for keeping HSPCs in the BM niche. We found that the expression of PAI-1, a downstream target of TGF-β signaling, was selectively augmented in niche-residing HSPCs. Functional inhibition of the TGF-β-PAI-1 signal increased MT1-MMP-dependent cellular motility, causing a detachment of HSPCs from the TGF-β-expressing niche cells, such as megakaryocytes. Furthermore, consistently high motility in PAI-1-deficient HSPCs was demonstrated by both a transwell migration assay and reciprocal transplantation experiments, indicating that intracellular, not extracellular, PAI-1 suppresses the motility of HSPCs, thereby causing them to stay in the niche. Mechanistically, intracellular PAI-1 inhibited the proteolytic activity of proprotein convertase Furin, diminishing MT1-MMP activity. This reduced expression of MT1-MMP in turn affected the expression levels of several adhesion/deadhesion molecules for determination of HSPC localization, such as CD44, VLA-4, and CXCR4, which then promoted the retention of HSPCs in the niche. Our findings open up a new field for the study of intracellular proteolysis as a regulatory mechanism of stem cell fate, which has the potential to improve clinical HSPC mobilization and transplantation protocols.
© 2017 by The American Society of Hematology.
Figures
Similar articles
-
Sphingosine-1-Phosphate Receptor-3 Supports Hematopoietic Stem and Progenitor Cell Residence Within the Bone Marrow Niche.Stem Cells. 2017 Apr;35(4):1040-1052. doi: 10.1002/stem.2556. Epub 2017 Jan 19. Stem Cells. 2017. PMID: 28026131 Free PMC article.
-
Blockade of plasminogen activator inhibitor-1 empties bone marrow niche sufficient for donor hematopoietic stem cell engraftment without myeloablative conditioning.Biochem Biophys Res Commun. 2019 Aug 20;516(2):500-505. doi: 10.1016/j.bbrc.2019.06.076. Epub 2019 Jun 20. Biochem Biophys Res Commun. 2019. PMID: 31230745
-
Evidence that a lipolytic enzyme--hematopoietic-specific phospholipase C-β2--promotes mobilization of hematopoietic stem cells by decreasing their lipid raft-mediated bone marrow retention and increasing the promobilizing effects of granulocytes.Leukemia. 2016 Apr;30(4):919-28. doi: 10.1038/leu.2015.315. Epub 2015 Nov 19. Leukemia. 2016. PMID: 26582648 Free PMC article.
-
MT1-MMP and RECK: opposite and essential roles in hematopoietic stem and progenitor cell retention and migration.J Mol Med (Berl). 2011 Dec;89(12):1167-74. doi: 10.1007/s00109-011-0792-9. Epub 2011 Aug 13. J Mol Med (Berl). 2011. PMID: 21842347 Review.
-
Novel techniques to quantitatively assess age-dependent alterations in biophysical properties of HSPCs and bone marrow niche.Exp Hematol. 2025 Feb;142:104686. doi: 10.1016/j.exphem.2024.104686. Epub 2024 Nov 28. Exp Hematol. 2025. PMID: 39613289 Review.
Cited by
-
Targeting of plasminogen activator inhibitor-1 activity promotes elimination of chronic myeloid leukemia stem cells.Haematologica. 2021 Feb 1;106(2):483-494. doi: 10.3324/haematol.2019.230227. Haematologica. 2021. PMID: 32001531 Free PMC article.
-
Damage of the Bone Marrow Stromal Precursors in Patients with Acute Leukemia at the Onset of the Disease and During Treatment.Int J Mol Sci. 2024 Dec 11;25(24):13285. doi: 10.3390/ijms252413285. Int J Mol Sci. 2024. PMID: 39769050 Free PMC article.
-
Protective effect of Huashi Baidu formula against AKI and active ingredients that target SphK1 and PAI-1.Chin Med. 2024 Nov 1;19(1):152. doi: 10.1186/s13020-024-01024-7. Chin Med. 2024. PMID: 39487526 Free PMC article.
-
Qigui-Yishen decoction delays renal fibrosis in mice with chronic kidney disease by regulating TM and PAI-1.Am J Transl Res. 2024 Jun 15;16(6):2358-2368. doi: 10.62347/GHOV4912. eCollection 2024. Am J Transl Res. 2024. PMID: 39006289 Free PMC article.
-
Perivascular Secretome Influences Hematopoietic Stem Cell Maintenance in a Gelatin Hydrogel.Ann Biomed Eng. 2021 Feb;49(2):780-792. doi: 10.1007/s10439-020-02602-0. Epub 2020 Sep 16. Ann Biomed Eng. 2021. PMID: 32939609 Free PMC article.
References
-
- Wilson A, Trumpp A. Bone-marrow haematopoietic-stem-cell niches. Nat Rev Immunol. 2006;6(2):93-106. - PubMed
-
- Wilson A, Oser GM, Jaworski M, et al. . Dormant and self-renewing hematopoietic stem cells and their niches. Ann N Y Acad Sci. 2007;1106(1):64-75. - PubMed
-
- Lapidot T, Petit I. Current understanding of stem cell mobilization: the roles of chemokines, proteolytic enzymes, adhesion molecules, cytokines, and stromal cells. Exp Hematol. 2002;30(9):973-981. - PubMed
-
- Lapid K, Glait-Santar C, Gur-Cohen S, et al. . Egress and Mobilization of Hematopoietic Stem and Progenitor Cells: A Dynamic Multi-facet Process. Cambridge, MA: Harvard Stem Cell Institute; 2012. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
