

Dissociable Effects on Birdsong of Androgen Signaling in Cortex-Like Brain Regions of Canaries
- PMID: 28821656
- PMCID: PMC5588457
- DOI: 10.1523/JNEUROSCI.3371-16.2017
Dissociable Effects on Birdsong of Androgen Signaling in Cortex-Like Brain Regions of Canaries
Abstract
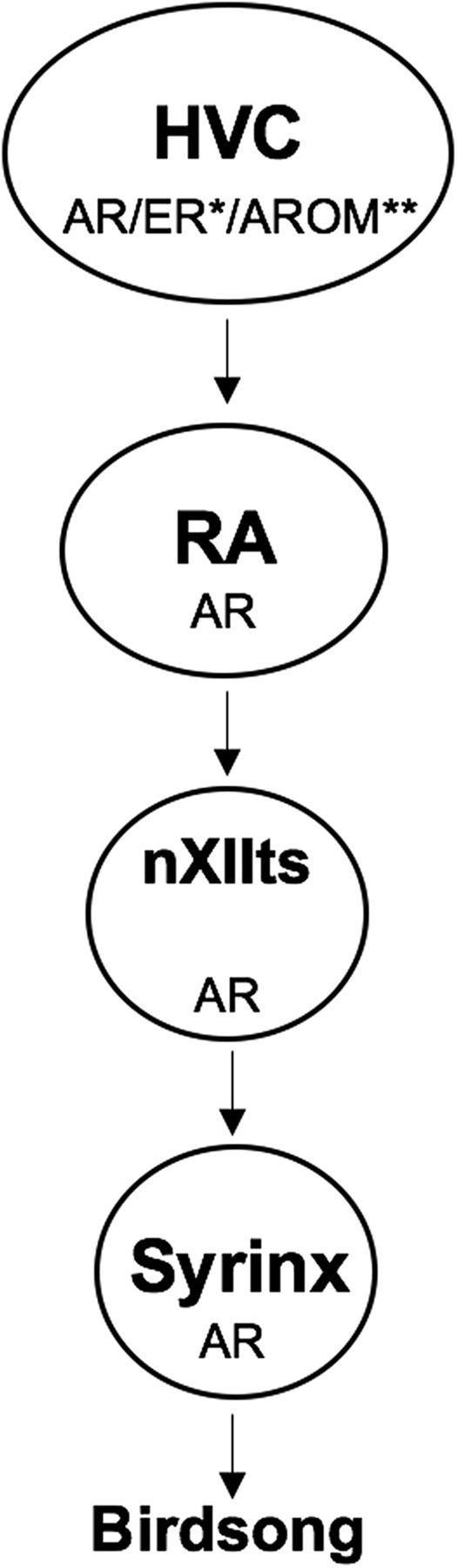
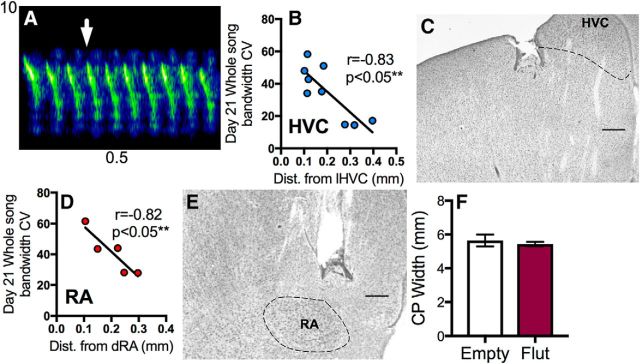
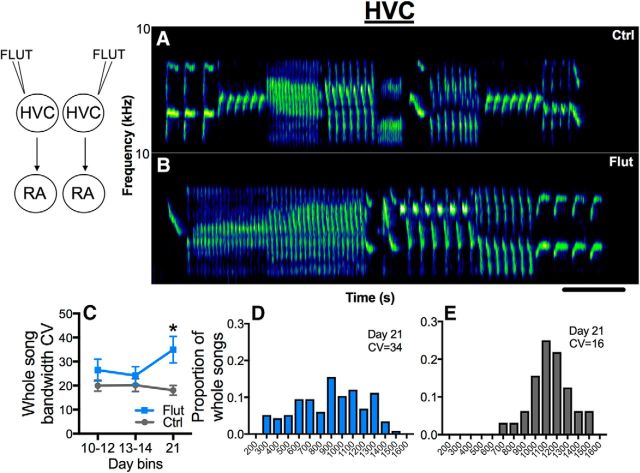
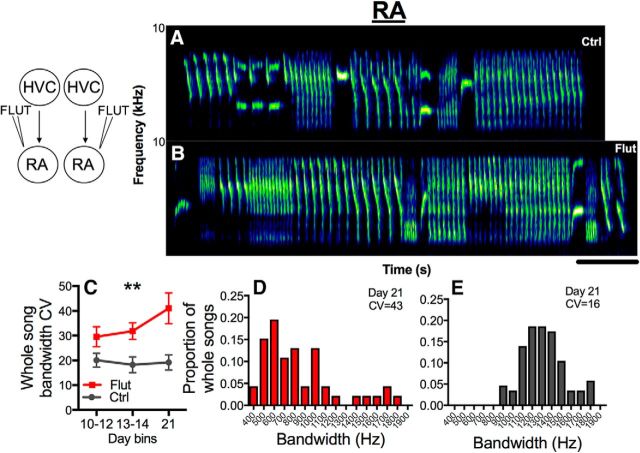
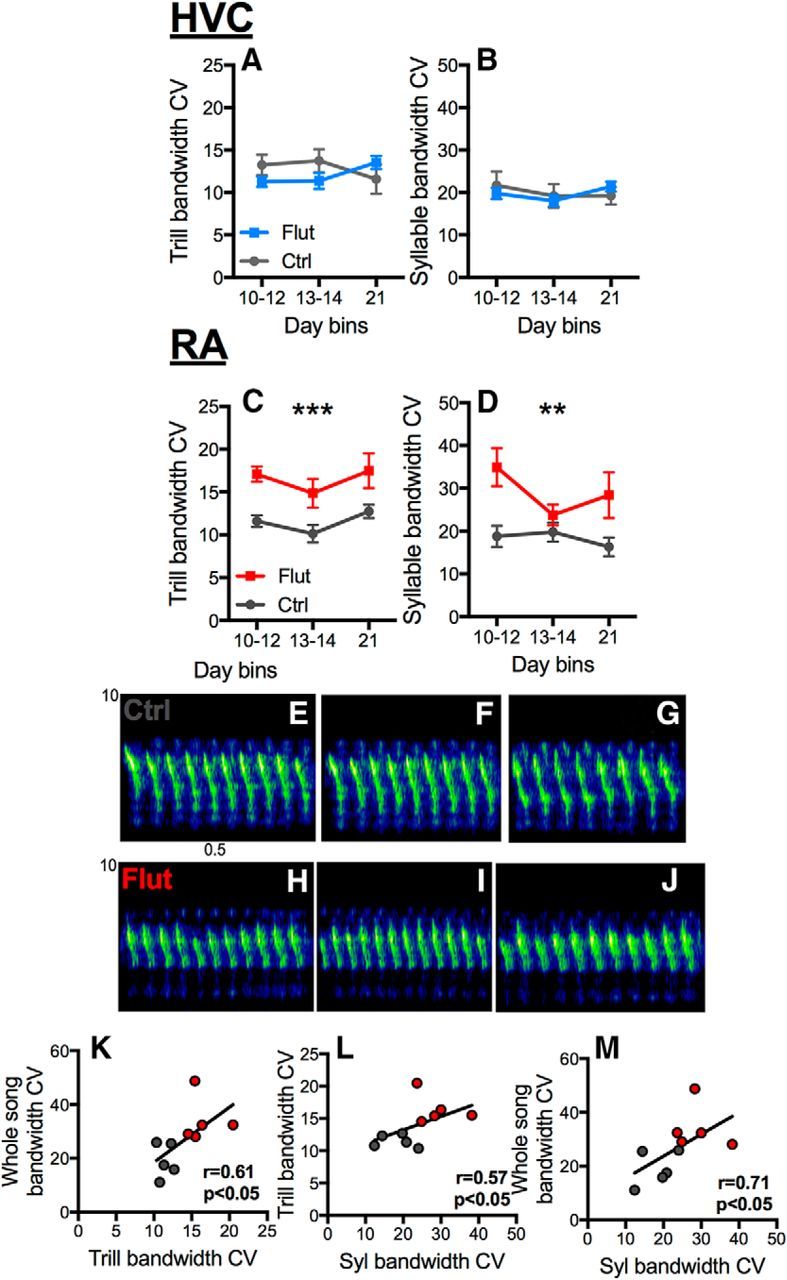
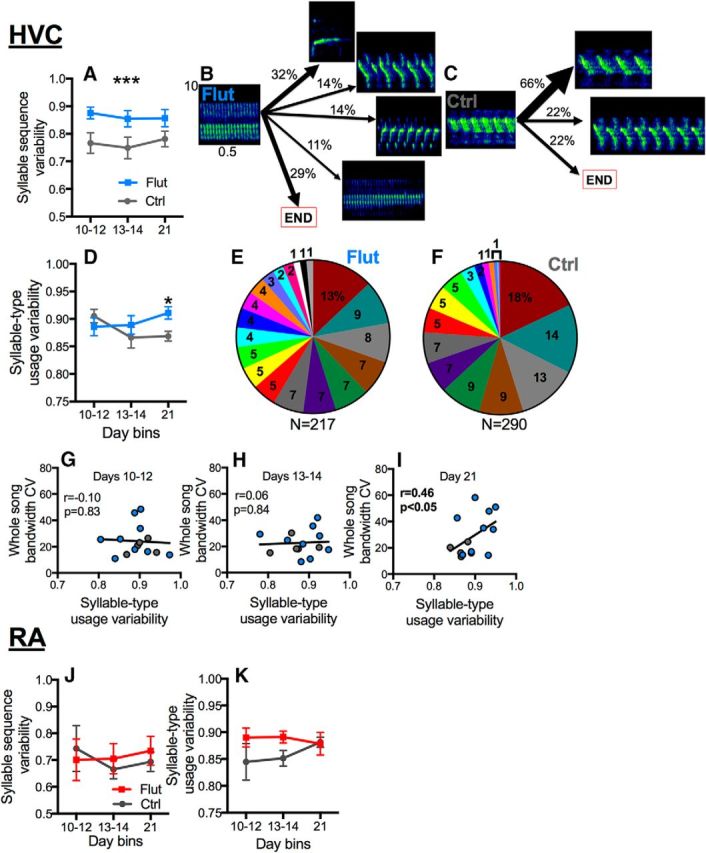
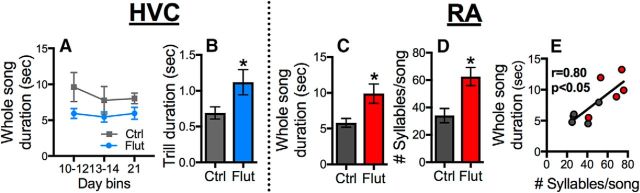
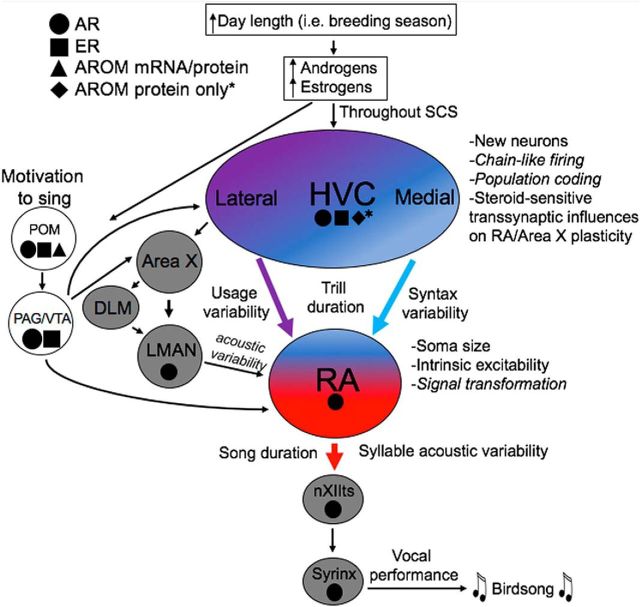
The neural basis of how learned vocalizations change during development and in adulthood represents a major challenge facing cognitive neuroscience. This plasticity in the degree to which learned vocalizations can change in both humans and songbirds is linked to the actions of sex steroid hormones during ontogeny but also in adulthood in the context of seasonal changes in birdsong. We investigated the role of steroid hormone signaling in the brain on distinct features of birdsong using adult male canaries (Serinus canaria), which show extensive seasonal vocal plasticity as adults. Specifically, we bilaterally implanted the potent androgen receptor antagonist flutamide in two key brain regions that control birdsong. We show that androgen signaling in the motor cortical-like brain region, the robust nucleus of the arcopallium (RA), controls syllable and trill bandwidth stereotypy, while not significantly affecting higher order features of song such syllable-type usage (i.e., how many times each syllable type is used) or syllable sequences. In contrast, androgen signaling in the premotor cortical-like brain region, HVC (proper name), controls song variability by increasing the variability of syllable-type usage and syllable sequences, while having no effect on syllable or trill bandwidth stereotypy. Other aspects of song, such as the duration of trills and the number of syllables per song, were also differentially affected by androgen signaling in HVC versus RA. These results implicate androgens in regulating distinct features of complex motor output in a precise and nonredundant manner.SIGNIFICANCE STATEMENT Vocal plasticity is linked to the actions of sex steroid hormones, but the precise mechanisms are unclear. We investigated this question in adult male canaries (Serinus canaria), which show extensive vocal plasticity throughout their life. We show that androgens in two cortex-like vocal control brain regions regulate distinct aspects of vocal plasticity. For example, in HVC (proper name), androgens regulate variability in syntax but not phonology, whereas androgens in the robust nucleus of the arcopallium (RA) regulate variability in phonology but not syntax. Temporal aspects of song were also differentially affected by androgen signaling in HVC versus RA. Thus, androgen signaling may reduce vocal plasticity by acting in a nonredundant and precise manner in the brain.
Keywords: androgens; birdsong; neuroendocrinology; steroid hormones; vocal plasticity.
Copyright © 2017 the authors 0270-6474/17/378612-13$15.00/0.
Figures
Similar articles
-
Androgen signaling in LMAN regulates song stereotypy in male canaries.Horm Behav. 2024 Sep;165:105611. doi: 10.1016/j.yhbeh.2024.105611. Epub 2024 Jul 31. Horm Behav. 2024. PMID: 39089160
-
Characteristics of song, brain-anatomy and blood androgen levels in spontaneously singing female canaries.Horm Behav. 2020 Jan;117:104614. doi: 10.1016/j.yhbeh.2019.104614. Epub 2019 Nov 1. Horm Behav. 2020. PMID: 31647922
-
Temperature Manipulation in Songbird Brain Implicates the Premotor Nucleus HVC in Birdsong Syntax.J Neurosci. 2017 Mar 8;37(10):2600-2611. doi: 10.1523/JNEUROSCI.1827-16.2017. Epub 2017 Feb 3. J Neurosci. 2017. PMID: 28159910 Free PMC article.
-
Sex differences in seasonal brain plasticity and the neuroendocrine regulation of vocal behavior in songbirds.Horm Behav. 2022 Jun;142:105160. doi: 10.1016/j.yhbeh.2022.105160. Epub 2022 Mar 30. Horm Behav. 2022. PMID: 35366412 Review.
-
How does testosterone act to regulate a multifaceted adaptive response? Lessons from studies of the avian song system.J Neuroendocrinol. 2020 Jan;32(1):e12793. doi: 10.1111/jne.12793. Epub 2019 Oct 2. J Neuroendocrinol. 2020. PMID: 31514252 Review.
Cited by
-
Androgen signaling in LMAN regulates song stereotypy in male canaries.Horm Behav. 2024 Sep;165:105611. doi: 10.1016/j.yhbeh.2024.105611. Epub 2024 Jul 31. Horm Behav. 2024. PMID: 39089160
-
Breaking Through the Bottleneck: Krogh's Principle in Behavioral Neuroendocrinology and the Potential of Gene Editing.Integr Comp Biol. 2023 Aug 23;63(2):428-443. doi: 10.1093/icb/icad068. Integr Comp Biol. 2023. PMID: 37312279 Free PMC article. Review.
-
A brain for all seasons: An in vivo MRI perspective on songbirds.J Exp Zool A Ecol Integr Physiol. 2022 Dec;337(9-10):967-984. doi: 10.1002/jez.2650. Epub 2022 Aug 21. J Exp Zool A Ecol Integr Physiol. 2022. PMID: 35989548 Free PMC article. Review.
-
Photoperiodism, testosterone and adult neurogenesis in canaries (Serinus canaria).J Neuroendocrinol. 2025 Jun;37(6):e13449. doi: 10.1111/jne.13449. Epub 2024 Sep 17. J Neuroendocrinol. 2025. PMID: 39286957 Review.
-
Anadara granosa shell powder improves the metabolism, testosterone level, and sound frequency of Pelung chickens.Vet World. 2021 Jun;14(6):1564-1571. doi: 10.14202/vetworld.2021.1564-1571. Epub 2021 Jun 18. Vet World. 2021. PMID: 34316204 Free PMC article.
References
-
- Alward BA, Madison FN, Gravley WT, Ball GF (2016b) Antagonism of syringeal androgen receptors reduces the quality of female-preferred male song in canaries. Anim Behav 119:201–212. 10.1016/j.anbehav.2016.07.010 - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources