

Drought and exogenous abscisic acid alter hydrogen peroxide accumulation and differentially regulate the expression of two maize RD22-like genes
- PMID: 28821770
- PMCID: PMC5562893
- DOI: 10.1038/s41598-017-08976-x
Drought and exogenous abscisic acid alter hydrogen peroxide accumulation and differentially regulate the expression of two maize RD22-like genes
Abstract
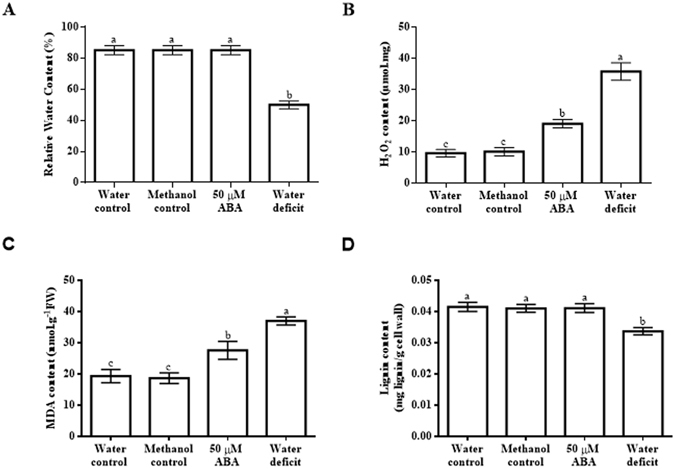
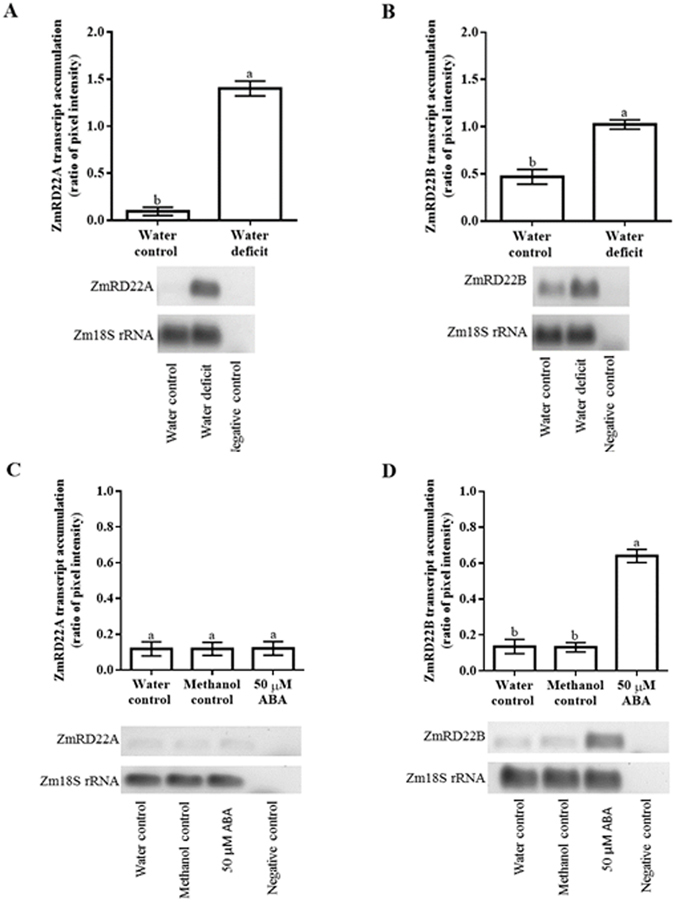
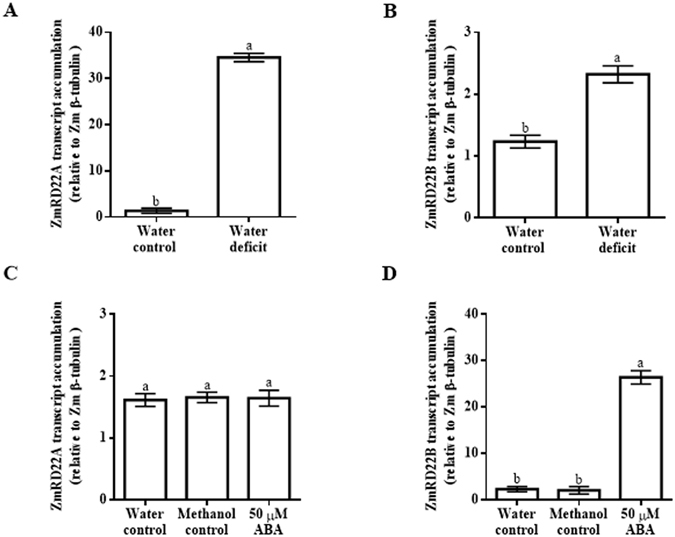
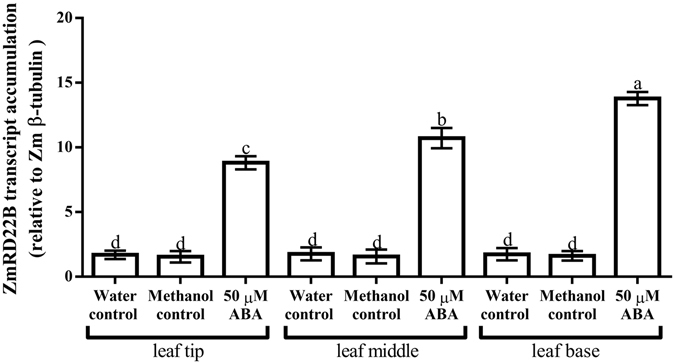
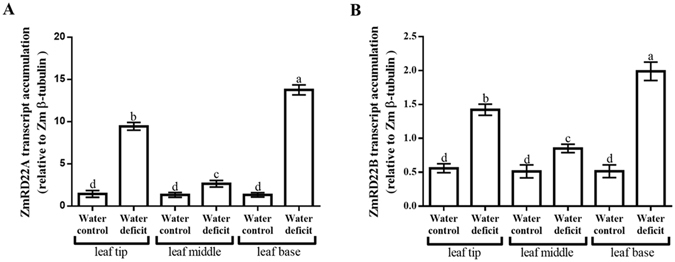
Increased biosynthesis of abscisic acid (ABA) occurs in plants in response to water deficit, which is mediated by changes in the levels of reactive oxygen species such as H2O2. Water deficit and ABA induce expression of some RD22-like proteins. This study aimed to evaluate the effect of water deficit and exogenous ABA (50 µM ABA applied every 24 hours for a total of 72 hours) on H2O2 content in Zea mays (maize) and to characterise genes encoding two putative maize RD22-like proteins (designated ZmRD22A and ZmRD22B). The expression profiles of the two putative maize RD22-like genes in response to water deficit and treatment with ABA were examined in leaves. In silico analyses showed that the maize RD22-like proteins share domain organisation with previously characterized RD22-like proteins. Both water deficit and exogenous ABA resulted in increased H2O2 content in leaves but the increase was more pronounced in response to water deficit than to exogenous ABA. Lignin content was not affected by exogenous ABA, whereas it was decreased by water deficit. Expression of both RD22-like genes was up-regulated by drought but the ZmRD22A gene was not influenced by exogenous ABA, whereas ZmRD22B was highly responsive to exogenous ABA.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
