

Distinct Neural Circuits for the Formation and Retrieval of Episodic Memories
- PMID: 28823555
- PMCID: PMC5586038
- DOI: 10.1016/j.cell.2017.07.013
Distinct Neural Circuits for the Formation and Retrieval of Episodic Memories
Abstract
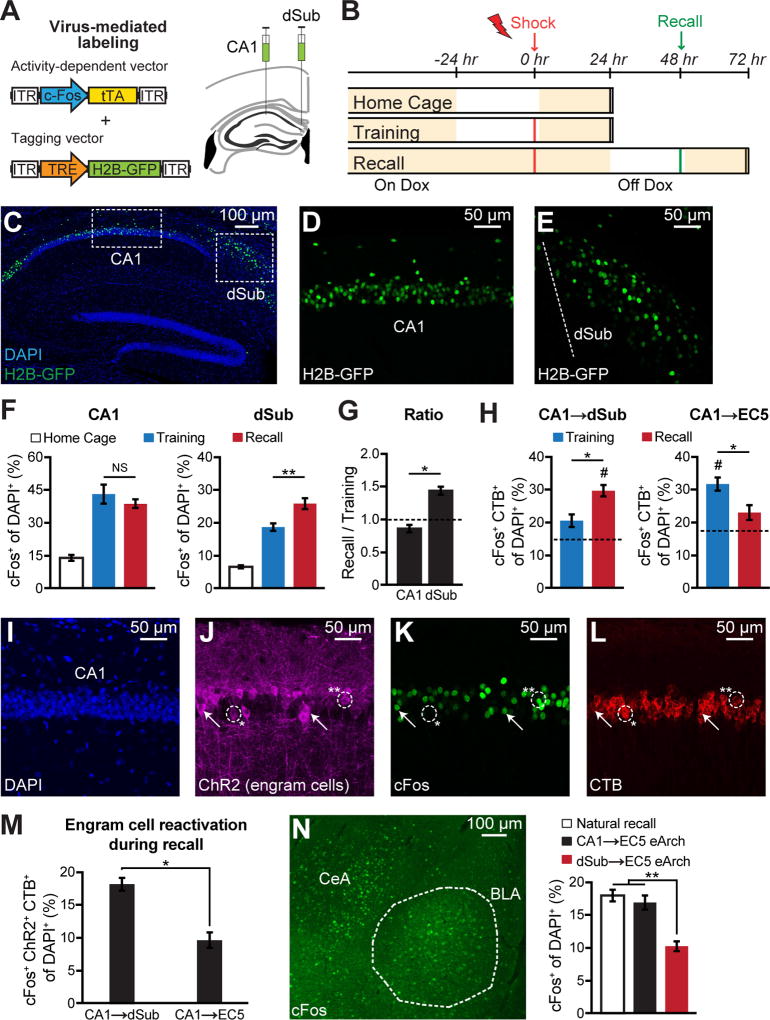
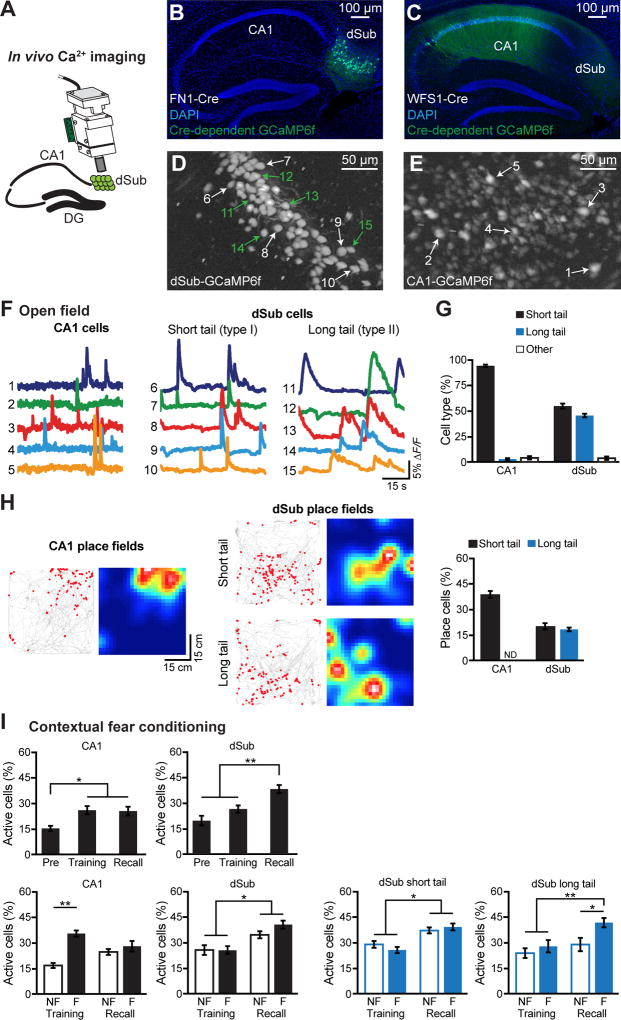
The formation and retrieval of a memory is thought to be accomplished by activation and reactivation, respectively, of the memory-holding cells (engram cells) by a common set of neural circuits, but this hypothesis has not been established. The medial temporal-lobe system is essential for the formation and retrieval of episodic memory for which individual hippocampal subfields and entorhinal cortex layers contribute by carrying out specific functions. One subfield whose function is poorly known is the subiculum. Here, we show that dorsal subiculum and the circuit, CA1 to dorsal subiculum to medial entorhinal cortex layer 5, play a crucial role selectively in the retrieval of episodic memories. Conversely, the direct CA1 to medial entorhinal cortex layer 5 circuit is essential specifically for memory formation. Our data suggest that the subiculum-containing detour loop is dedicated to meet the requirements associated with recall such as rapid memory updating and retrieval-driven instinctive fear responses.
Keywords: Subiculum; entorhinal cortex; episodic memory; hippocampus; mammillary bodies; memory formation; memory retrieval; memory updating; neural circuits; stress hormone.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
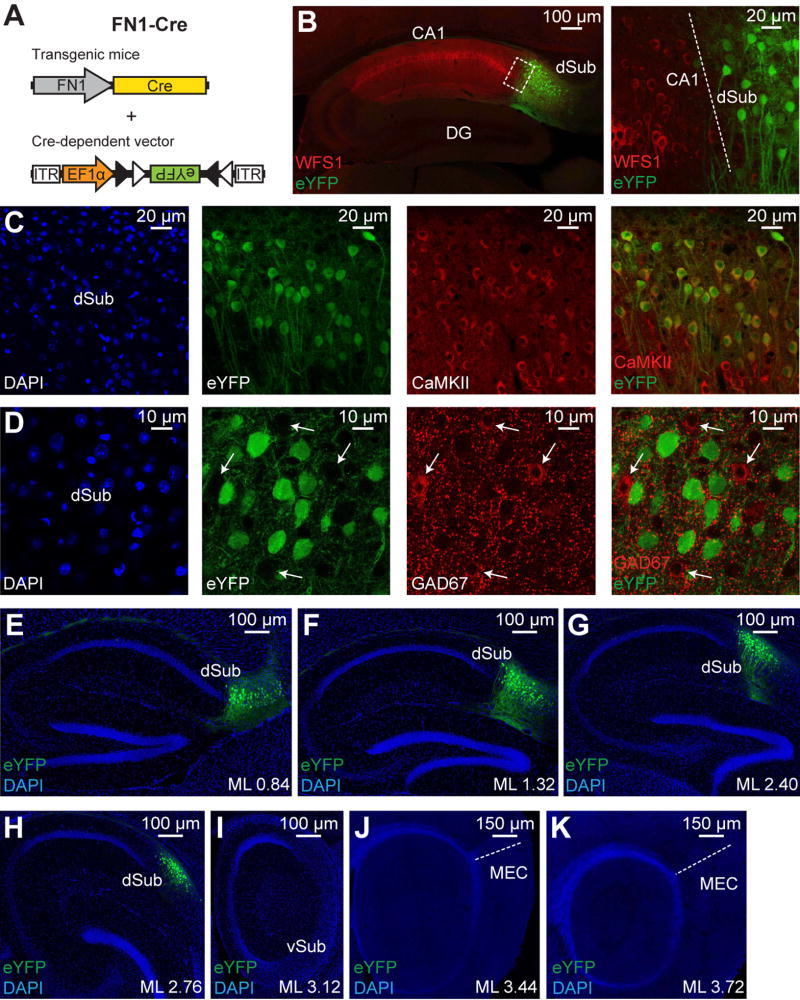
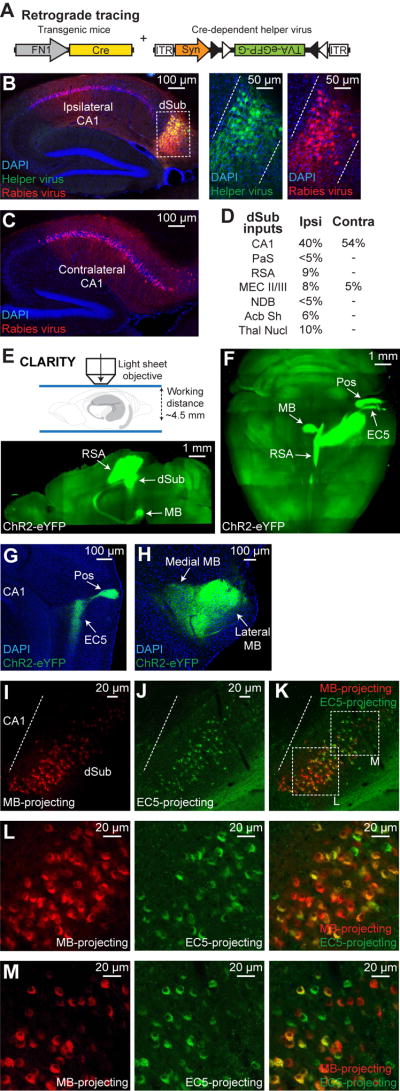
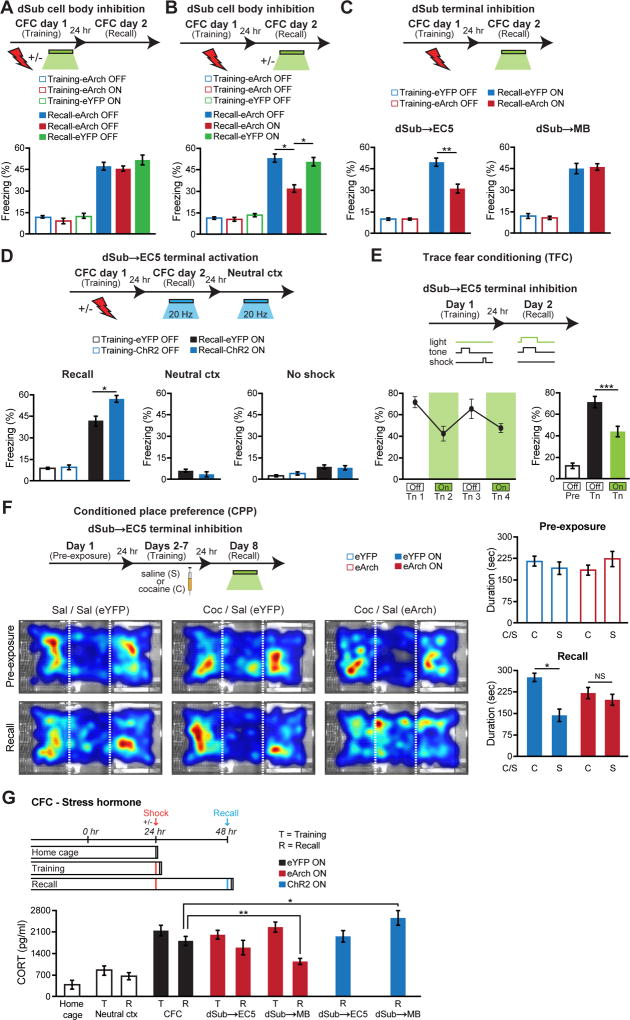
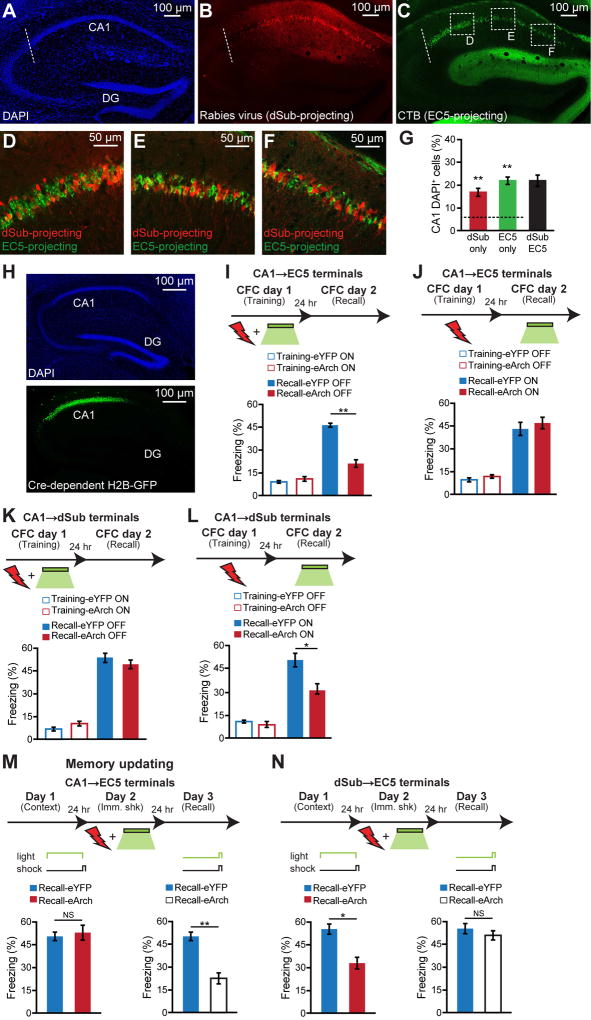
Comment in
-
Learning and memory: Memories take the sub-way.Nat Rev Neurosci. 2017 Sep 19;18(10):571. doi: 10.1038/nrn.2017.117. Nat Rev Neurosci. 2017. PMID: 28924255 No abstract available.
-
Memory Retrieval: Taking the Route via Subiculum.Curr Biol. 2017 Nov 20;27(22):R1225-R1227. doi: 10.1016/j.cub.2017.09.042. Curr Biol. 2017. PMID: 29161563
References
-
- Bourque CW, Oliet SH, Kirkpatrick K, Richard D, Fisher TE. Extrinsic and intrinsic modulatory mechanisms involved in regulating the electrical activity of supraoptic neurons. Ann. N. Y. Acad. Sci. 1993;689:512–519. - PubMed
-
- Ding SL. Comparative anatomy of the prosubiculum, subiculum, presubiculum, postsubiculum, and parasubiculum in human, monkey, and rodent. J. Comp. Neurol. 2013;521:4145–4162. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
