

Midbrain-Hindbrain Boundary Morphogenesis: At the Intersection of Wnt and Fgf Signaling
- PMID: 28824384
- PMCID: PMC5541008
- DOI: 10.3389/fnana.2017.00064
Midbrain-Hindbrain Boundary Morphogenesis: At the Intersection of Wnt and Fgf Signaling
Abstract
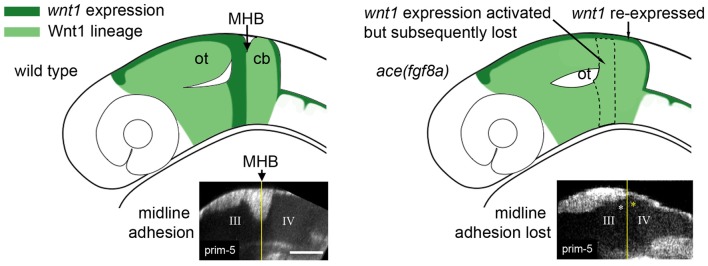
A constriction in the neural tube at the junction of the midbrain and hindbrain is a conserved feature of vertebrate embryos. The constriction is a defining feature of the midbrain-hindbrain boundary (MHB), a signaling center that patterns the adjacent midbrain and rostral hindbrain and forms at the junction of two gene expression domains in the early neural plate: an anterior otx2/wnt1 positive domain and a posterior gbx/fgf8 positive domain. otx2 and gbx genes encode mutually repressive transcription factors that create a lineage restriction boundary at their expression interface. Wnt and Fgf genes form a mutually dependent feedback system that maintains their expression domains on the otx2 or gbx side of the boundary, respectively. Constriction morphogenesis occurs after these conserved gene expression domains are established and while their mutual interactions maintain their expression pattern; consequently, mutant studies in zebrafish have led to the suggestion that constriction morphogenesis should be considered a unique phase of MHB development. We analyzed MHB morphogenesis in fgf8 loss of function zebrafish embryos using a reporter driven by the conserved wnt1 enhancer to visualize anterior boundary cells. We found that fgf8 loss of function results in a re-activation of wnt1 reporter expression posterior to the boundary simultaneous with an inactivation of the wnt1 reporter in the anterior boundary cells, and that these events correlate with relaxation of the boundary constriction. In consideration of other results that correlate the boundary constriction with Wnt and Fgf expression, we propose that the maintenance of an active Wnt-Fgf feedback loop is a key factor in driving the morphogenesis of the MHB constriction.
Keywords: Fgf; MHB; Wnt; constriction morphogenesis; image analysis; mes/r1; two-photon fluorescence; zebrafish.
Figures





References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous