

DltX of Bacillus thuringiensis Is Essential for D-Alanylation of Teichoic Acids and Resistance to Antimicrobial Response in Insects
- PMID: 28824570
- PMCID: PMC5541007
- DOI: 10.3389/fmicb.2017.01437
DltX of Bacillus thuringiensis Is Essential for D-Alanylation of Teichoic Acids and Resistance to Antimicrobial Response in Insects
Abstract
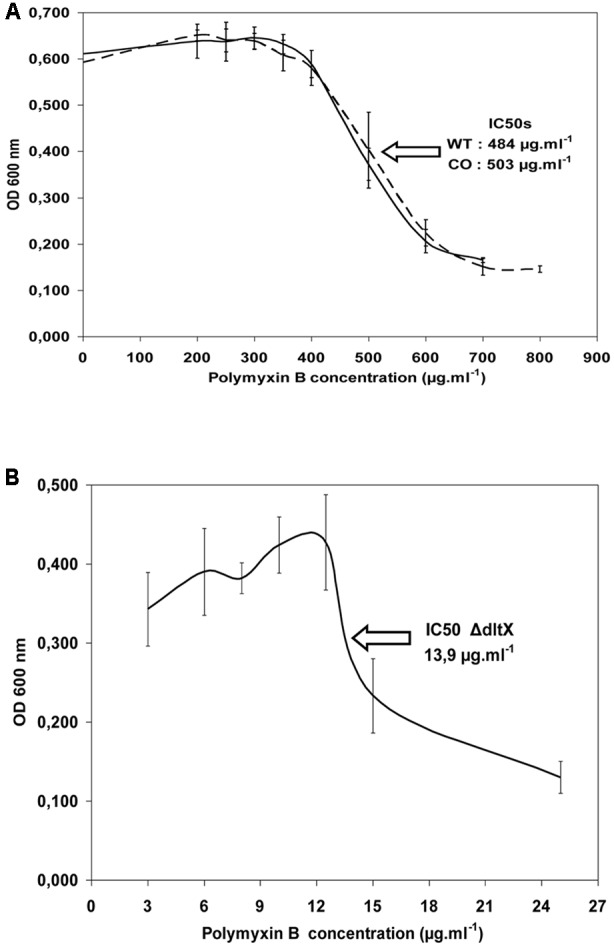
The dlt operon of Gram-positive bacteria is required for the incorporation of D-alanine esters into cell wall-associated teichoic acids (TAs). Addition of D-alanine to TAs reduces the negative charge of the cell envelope thereby preventing cationic antimicrobial peptides (CAMPs) from reaching their target of action on the bacterial surface. In most gram-positive bacteria, this operon consists of five genes dltXABCD but the involvement of the first ORF (dltX) encoding a small protein of unknown function, has never been investigated. The aim of this study was to establish whether this protein is involved in the D-alanylation process in Bacillus thuringiensis. We, therefore constructed an in frame deletion mutant of dltX, without affecting the expression of the other genes of the operon. The growth characteristics of the dltX mutant and those of the wild type strain were similar under standard in vitro conditions. However, disruption of dltX drastically impaired the resistance of B. thuringiensis to CAMPs and significantly attenuated its virulence in two insect species. Moreover, high-performance liquid chromatography studies showed that the dltX mutant was devoid of D-alanine, and electrophoretic mobility measurements indicated that the cells carried a higher negative surface charge. Scanning electron microscopy experiments showed morphological alterations of these mutant bacteria, suggesting that depletion of D-alanine from TAs affects cell wall structure. Our findings suggest that DltX is essential for the incorporation of D-alanyl esters into TAs. Therefore, DltX plays a direct role in the resistance to CAMPs, thus contributing to the survival of B. thuringiensis in insects. To our knowledge, this work is the first report examining the involvement of dltX in the D-alanylation of TAs.
Keywords: B. thuringiensis; D-alanylation; antimicrobial peptides; dltX; innate immunity; insects; virulence.
Figures







References
-
- Abi Khattar Z., Rejasse A., Destoumieux-Garzón D., Escoubas J. M., Sanchis V., Lereclus D., et al. (2009). The dlt operon of Bacillus cereus is required for resistance to cationic antimicrobial peptides and for virulence in insects. J. Bacteriol. 191 7063–7073. 10.1128/JB.00892-09 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases