

Trypanosoma Infection Favors Brucella Elimination via IL-12/IFNγ-Dependent Pathways
- PMID: 28824630
- PMCID: PMC5534484
- DOI: 10.3389/fimmu.2017.00903
Trypanosoma Infection Favors Brucella Elimination via IL-12/IFNγ-Dependent Pathways
Abstract
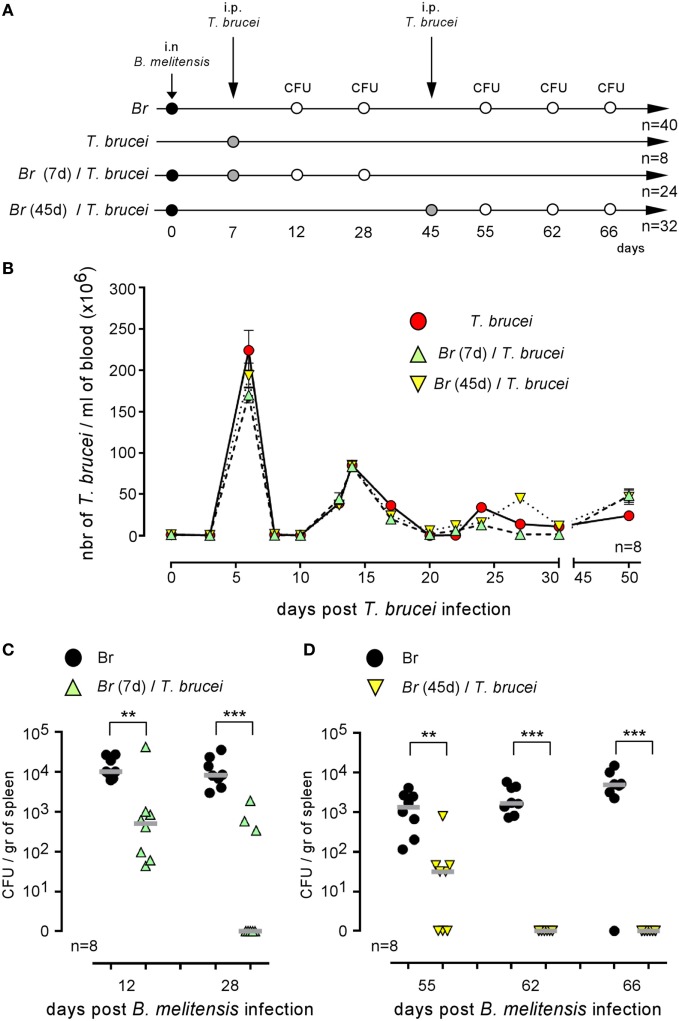
This study develops an original co-infection model in mice using Brucella melitensis, the most frequent cause of human brucellosis, and Trypanosoma brucei, the agent of African trypanosomiasis. Although the immunosuppressive effects of T. brucei in natural hosts and mice models are well established, we observed that the injection of T. brucei in mice chronically infected with B. melitensis induces a drastic reduction in the number of B. melitensis in the spleen, the main reservoir of the infection. Similar results are obtained with Brucella abortus- and Brucella suis-infected mice and B. melitensis-infected mice co-infected with Trypanosoma cruzi, demonstrating that this phenomenon is not due to antigenic cross-reactivity. Comparison of co-infected wild-type and genetically deficient mice showed that Brucella elimination required functional IL-12p35/IFNγ signaling pathways and the presence of CD4+ T cells. However, the impact of wild type and an attenuated mutant of T. brucei on B. melitensis were similar, suggesting that a chronic intense inflammatory reaction is not required to eliminate B. melitensis. Finally, we also tested the impact of T. brucei infection on the course of Mycobacterium tuberculosis infection. Although T. brucei strongly increases the frequency of IFNγ+CD4+ T cells, it does not ameliorate the control of M. tuberculosis infection, suggesting that it is not controlled by the same effector mechanisms as Brucella. Thus, whereas T. brucei infections are commonly viewed as immunosuppressive and pathogenic, our data suggest that these parasites can specifically affect the immune control of Brucella infection, with benefits for the host.
Keywords: Brucella abortus; Brucella melitensis; Mycobacterium tuberculosis; Trypanosoma brucei brucei; Trypanosoma cruzi; brucellosis; infection control.
Figures







References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials