

Knock-Down of the IFR1 Protein Perturbs the Homeostasis of Reactive Electrophile Species and Boosts Photosynthetic Hydrogen Production in Chlamydomonas reinhardtii
- PMID: 28824682
- PMCID: PMC5540887
- DOI: 10.3389/fpls.2017.01347
Knock-Down of the IFR1 Protein Perturbs the Homeostasis of Reactive Electrophile Species and Boosts Photosynthetic Hydrogen Production in Chlamydomonas reinhardtii
Abstract
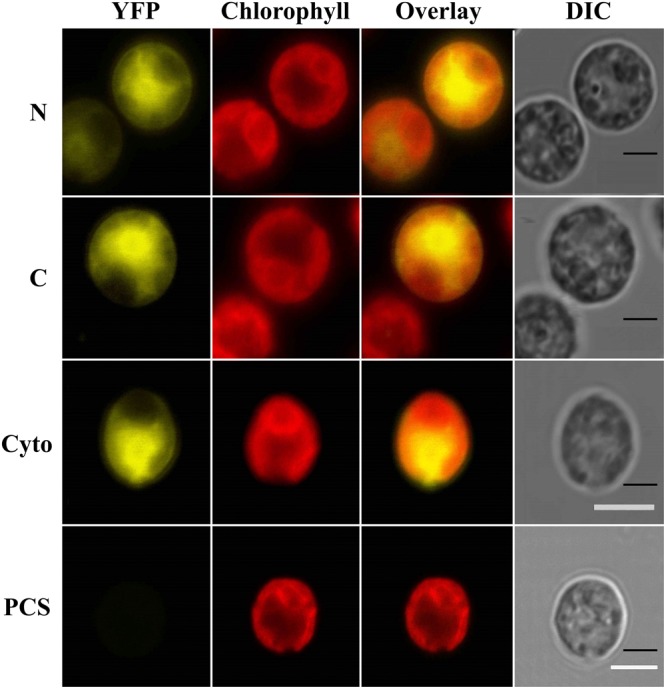
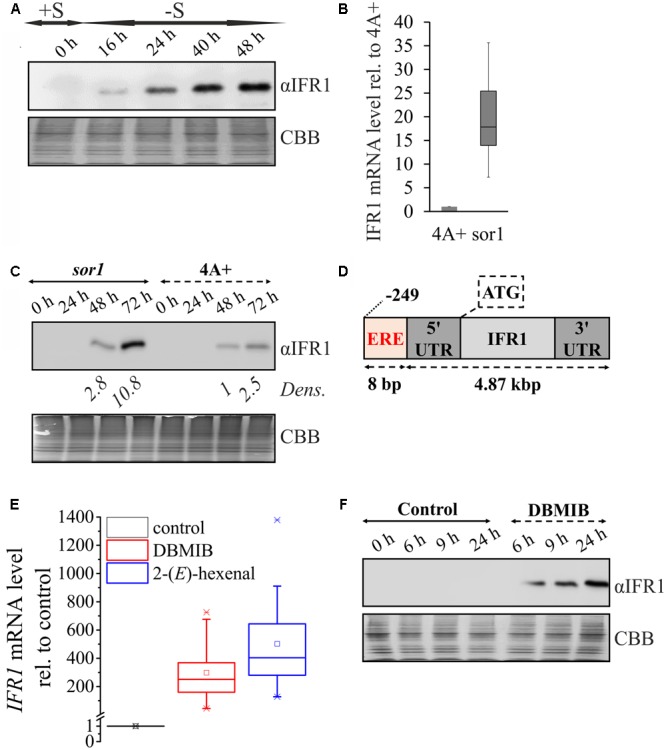
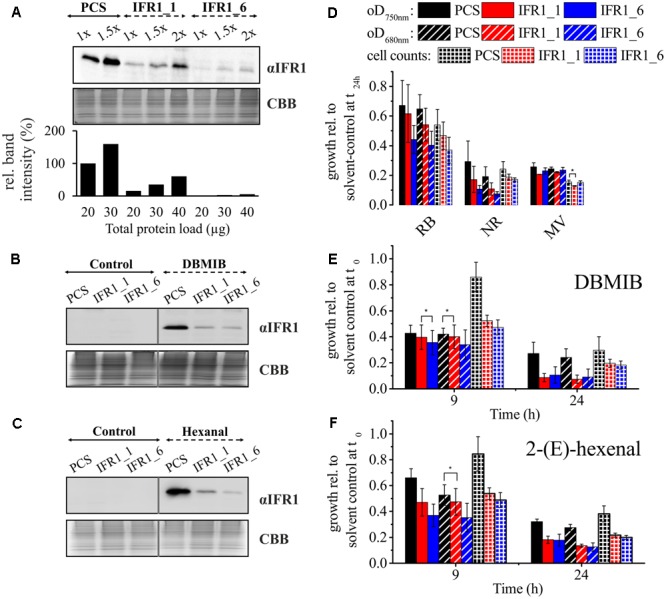
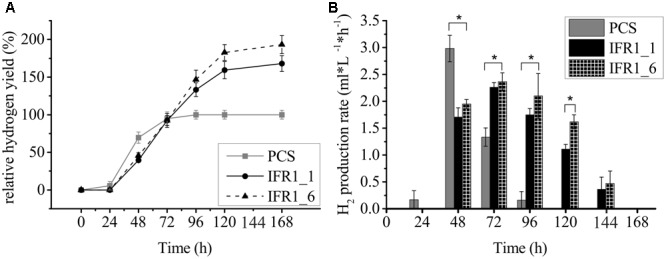
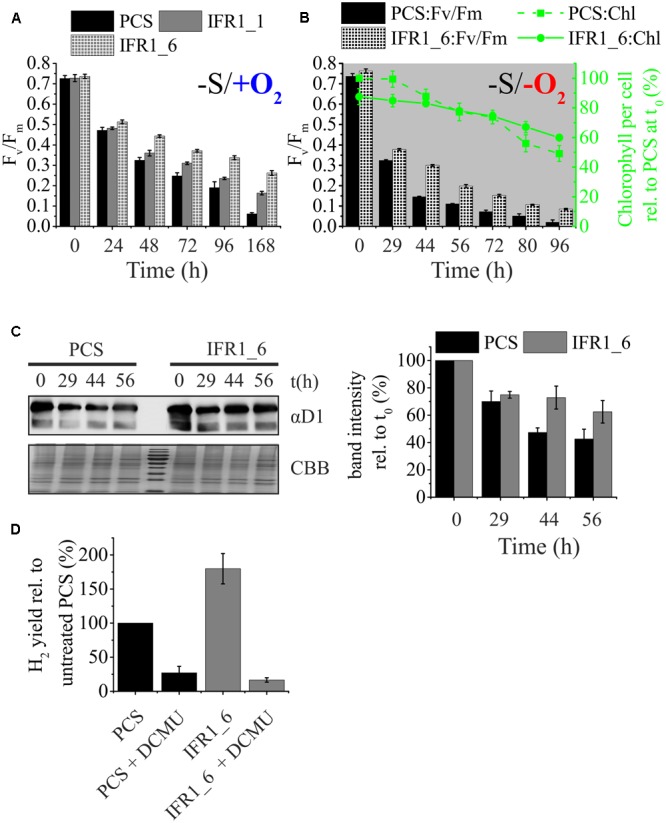
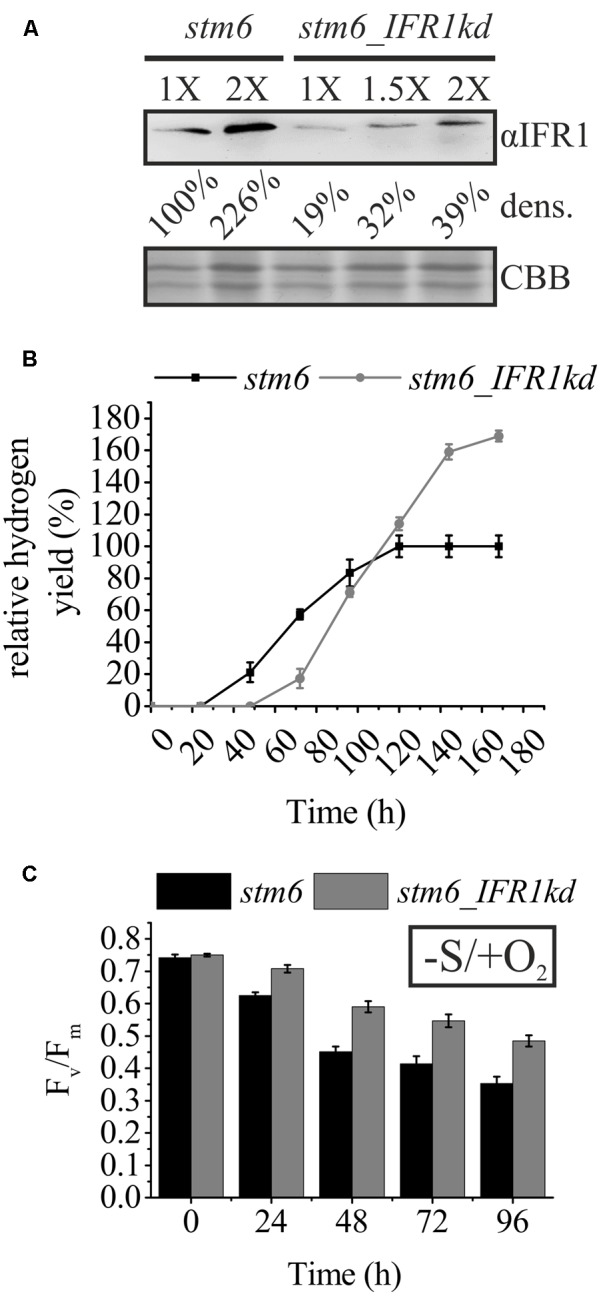
The protein superfamily of short-chain dehydrogenases/reductases (SDR), including members of the atypical type (aSDR), covers a huge range of catalyzed reactions and in vivo substrates. This superfamily also comprises isoflavone reductase-like (IRL) proteins, which are aSDRs highly homologous to isoflavone reductases from leguminous plants. The molecular function of IRLs in non-leguminous plants and green microalgae has not been identified as yet, but several lines of evidence point at their implication in reactive oxygen species homeostasis. The Chlamydomonas reinhardtii IRL protein IFR1 was identified in a previous study, analyzing the transcriptomic changes occurring during the acclimation to sulfur deprivation and anaerobiosis, a condition that triggers photobiological hydrogen production in this microalgae. Accumulation of the cytosolic IFR1 protein is induced by sulfur limitation as well as by the exposure of C. reinhardtii cells to reactive electrophile species (RES) such as reactive carbonyls. The latter has not been described for IRL proteins before. Over-accumulation of IFR1 in the singlet oxygen response 1 (sor1) mutant together with the presence of an electrophile response element, known to be required for SOR1-dependent gene activation as a response to RES, in the promoter of IFR1, indicate that IFR1 expression is controlled by the SOR1-dependent pathway. An implication of IFR1 into RES homeostasis, is further implied by a knock-down of IFR1, which results in a diminished tolerance toward RES. Intriguingly, IFR1 knock-down has a positive effect on photosystem II (PSII) stability under sulfur-deprived conditions used to trigger photobiological hydrogen production, by reducing PSII-dependent oxygen evolution, in C. reinhardtii. Reduced PSII photoinhibition in IFR1 knock-down strains prolongs the hydrogen production phase resulting in an almost doubled final hydrogen yield compared to the parental strain. Finally, IFR1 knock-down could be successfully used to further increase hydrogen yields of the high hydrogen-producing mutant stm6, demonstrating that IFR1 is a promising target for genetic engineering approaches aiming at an increased hydrogen production capacity of C. reinhardtii cells.
Keywords: Chlamydomonas reinhardtii; isoflavone reductase-like proteins; photobiological hydrogen production; reactive electrophile species; short-chain dehydrogenases/reductases; singlet oxygen response 1 (sor1).
Figures
References
-
- Annamalai J., Nallamuthu T. (2014). Antioxidant potential phytochemicals from methanol extract of Chlorella vulgaris and Chlamydomonas reinhardtii. J. Algal Biomass Util. 5 60–67.
-
- Antal T. K., Krendeleva T. E., Laurinavichene T. V., Makarova V. V., Ghirardi M. L., Rubin A. B., et al. (2003). The dependence of algal H2 production on Photosystem II and O2 consumption activities in sulfur-deprived Chlamydomonas reinhardtii cells. Biochim. Biophys. Acta 1607 153–160. 10.1016/j.bbabio.2003.09.008 - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous