

Post-discharge hyperpolarization is an endogenous modulatory factor limiting input from fast-conducting nociceptors (AHTMRs)
- PMID: 28825337
- PMCID: PMC5570122
- DOI: 10.1177/1744806917726255
Post-discharge hyperpolarization is an endogenous modulatory factor limiting input from fast-conducting nociceptors (AHTMRs)
Abstract
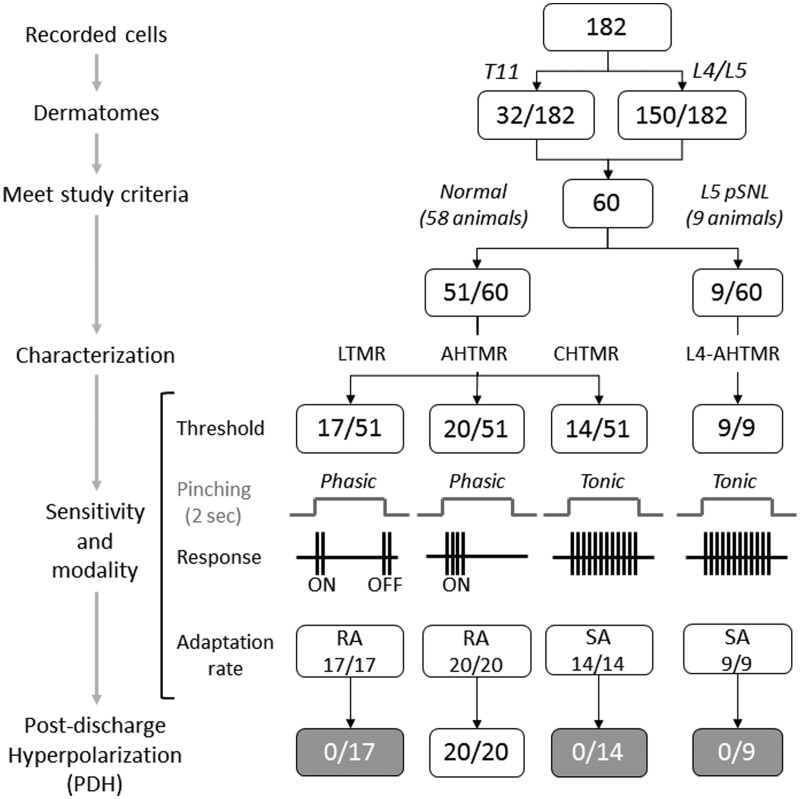
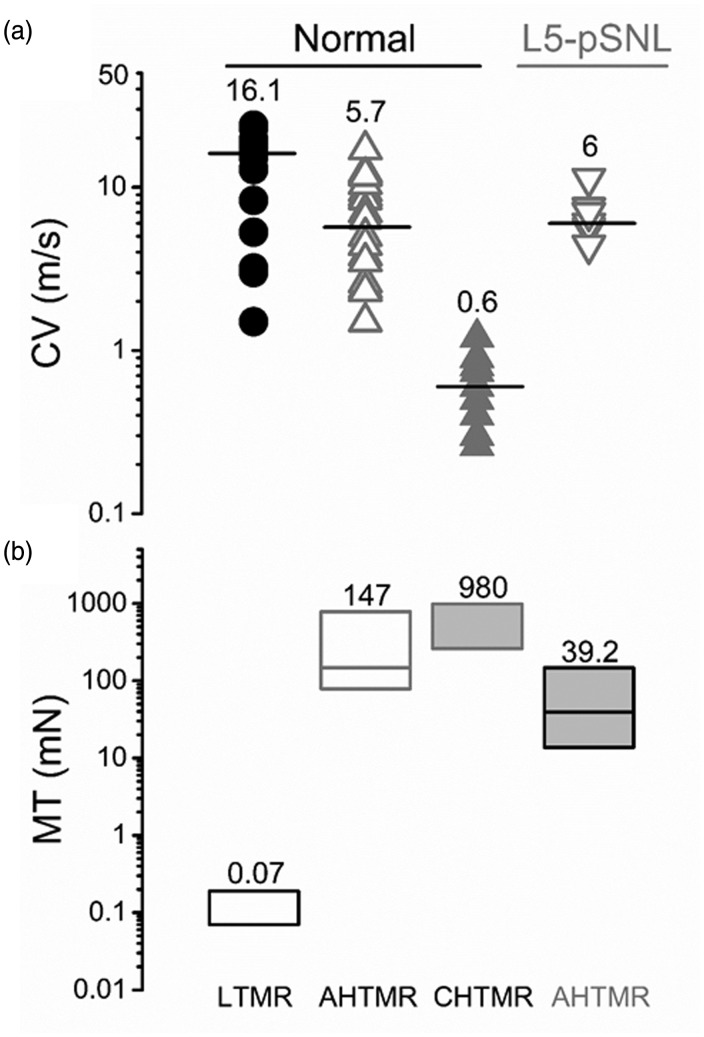
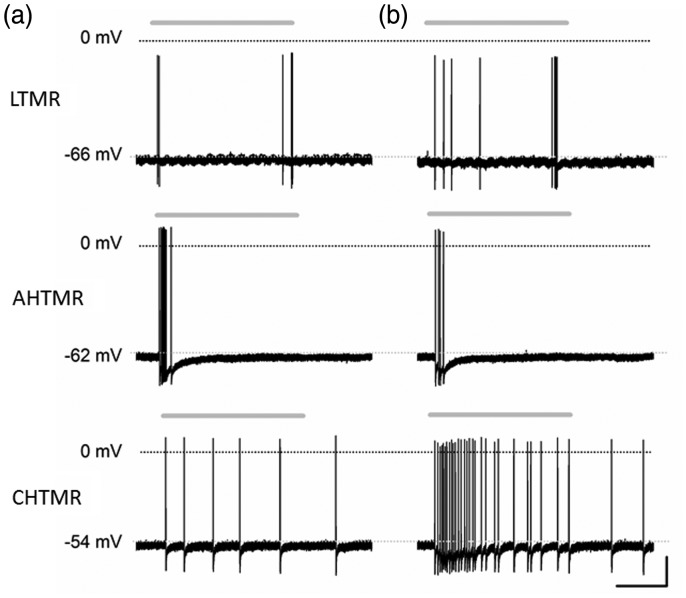
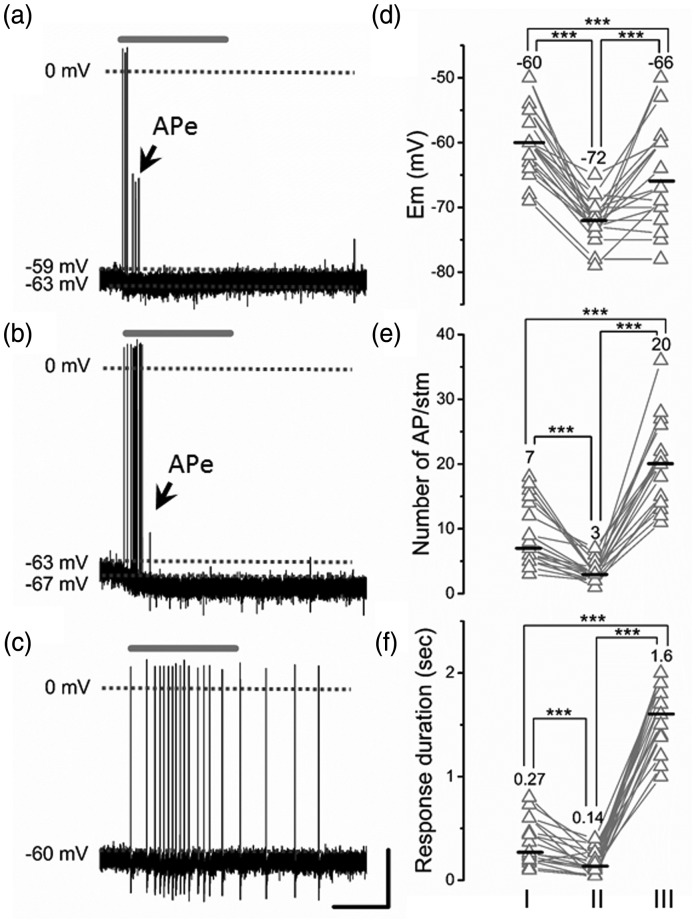
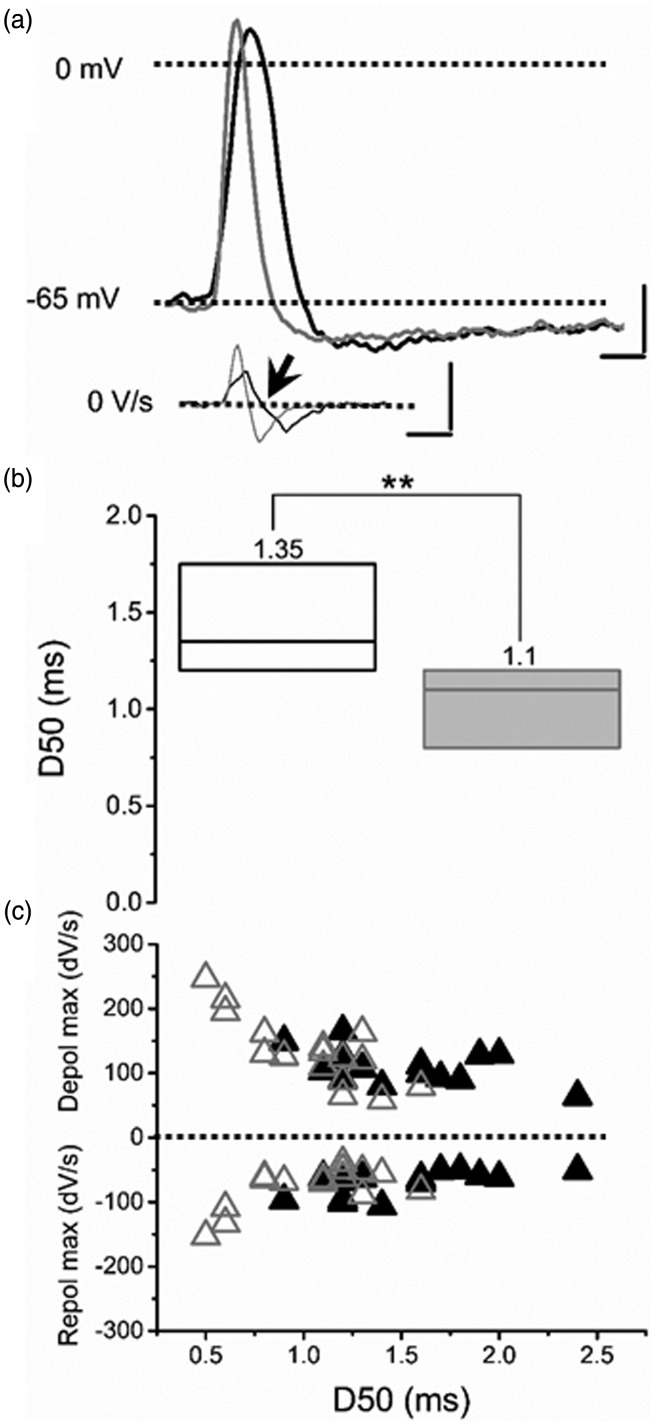
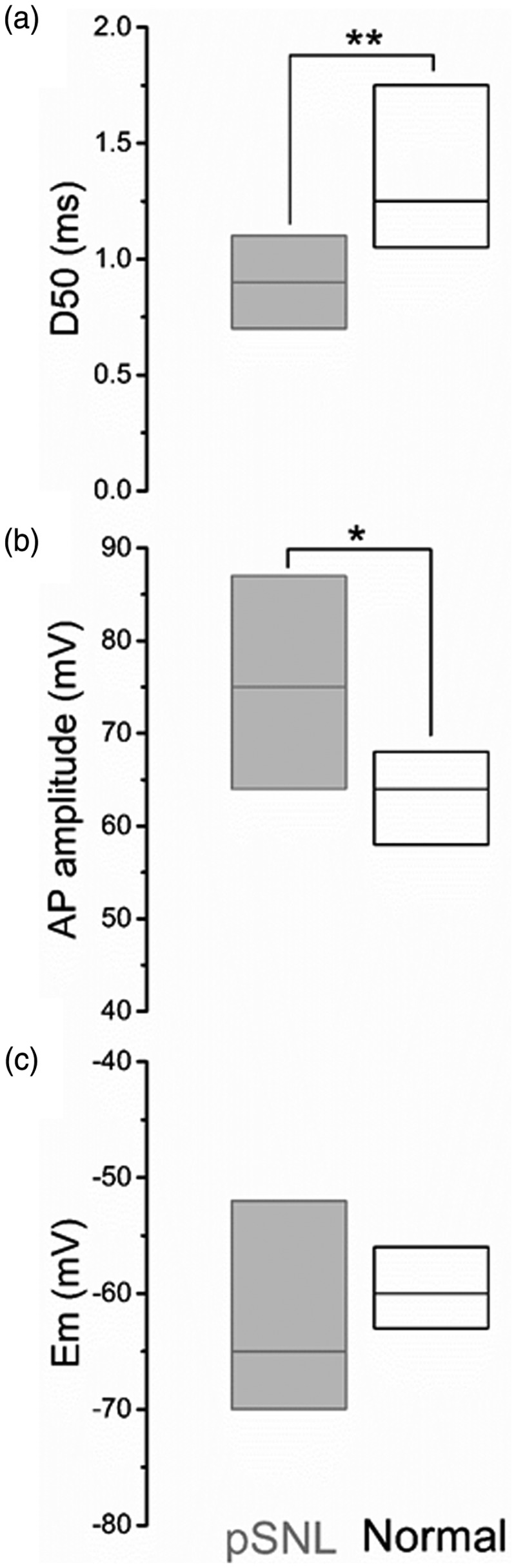
Peripheral somatosensory neurons are frequently exposed to mechanical forces. Strong stimuli result in neuronal activation of high-threshold mechanosensory afferent neurons, even in the absence of tissue damage. Among these neurons, fast-conducting nociceptors (A-fiber high-threshold mechanoreceptors (AHTMRs)) are normally resistant to sustained activation, transiently encoding the mechanical stimulus intensity but not its full duration. This rapidly adapting response seems to depend on changes in the electrical excitability of the membrane of these afferent neurons during sustained stimulation, a restraint mechanism that disappears following sensitization. Here, we examine the mechanism by which strong peripheral activation of mechanoreceptors elicits this control process in the absence of tissue injury and temporally silences afferent neurons despite ongoing stimulation. To study this, mechanoreceptors in Sprague-Dawley rats were accessed at the soma in the dorsal root ganglia from T11 and L4/L5. Neuronal classification was performed using receptive field characteristics and passive and active electrical properties. Sustained mechanical nociceptive stimulation in the absence of tissue damage of AHTMRs induces a rapid membrane hyperpolarization and a period of reduced responsiveness to the stimuli. Moreover, this phenomenon appears to be unique to this subset of afferent neurons and is absent in slow-conducting C-mechanonociceptors (C-fiber high-threshold mechanoreceptors) and rapidly adapting fast-conducting low-threshold mechanoreceptors. Furthermore, this mechanism for rapid adaptation and reducing ongoing input is ablated by repeated strong stimuli and in sensitized AHTMRs after chronic neuropathic injury. Further studies to understand the underling molecular mechanisms behind this phenomenon and their modulation during the development of pathological conditions may provide new targets to control nociceptive hyperexcitability and chronic pain.
Keywords: Primary sensory neurons; in vivo electrophysiology; membrane hyperpolarization.
Figures
References
-
- Andrew D, Greenspan JD. Peripheral coding of tonic mechanical cutaneous pain: comparison of nociceptor activity in rat and human psychophysics. J Neurophysiol 1999; 82: 2641–2648. - PubMed
-
- Boada MD. Relationship between electrophysiological signature and defined sensory modality of trigeminal ganglion neurons in vivo. J Neurophysiol 2013; 109: 749–757. DOI: 10.1152/jn.00693.2012. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
