

RING-Between-RING E3 Ligases: Emerging Themes amid the Variations
- PMID: 28827147
- PMCID: PMC5675740
- DOI: 10.1016/j.jmb.2017.08.008
RING-Between-RING E3 Ligases: Emerging Themes amid the Variations
Abstract
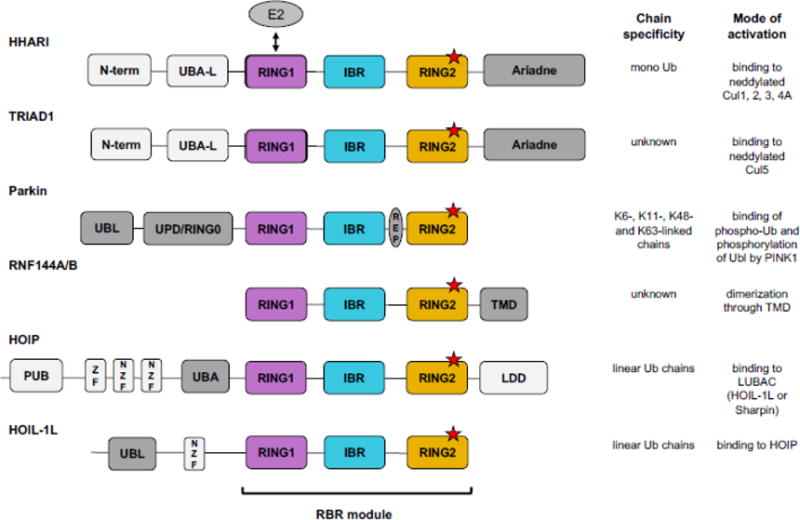
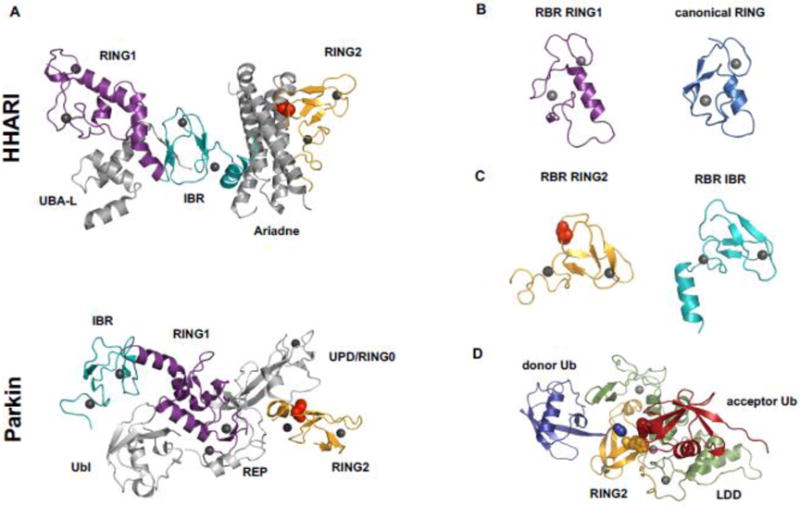
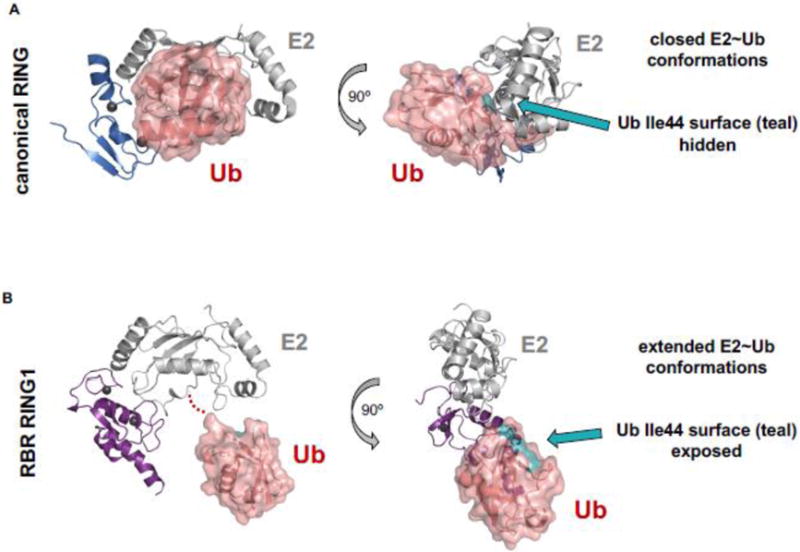
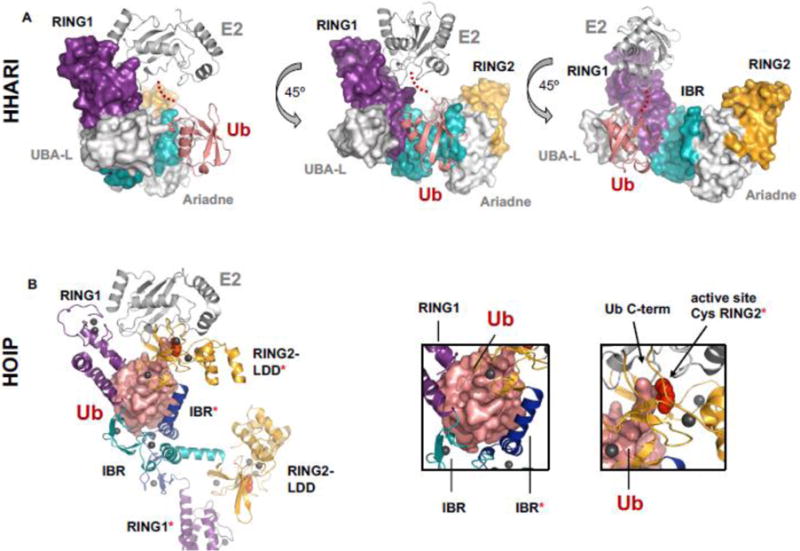
Covalent, reversible, post-translational modification of cellular proteins with the small modifier, ubiquitin (Ub), regulates virtually every known cellular process in eukaryotes. The process is carried out by a trio of enzymes: a Ub-activating (E1) enzyme, a Ub-conjugating (E2) enzyme, and a Ub ligase (E3) enzyme. RING-in-Between-RING (RBR) E3s constitute one of three classes of E3 ligases and are defined by a RING-HECT-hybrid mechanism that utilizes a E2-binding RING domain and a second domain (called RING2) that contains an active site Cys required for the formation of an obligatory E3~Ub intermediate. Albeit a small class, RBR E3s in humans regulate diverse cellular process. This review focuses on non-Parkin members such as HOIP/HOIL-1L (the only E3s known to generate linear Ub chains), HHARI and TRIAD1, both of which have been recently demonstrated to work together with Cullin RING E3 ligases. We provide a brief historical background and highlight, summarize, and discuss recent developments in the young field of RBR E3s. Insights reviewed here include new understandings of the RBR Ub-transfer mechanism, specifically the role of RING1 and various Ub-binding sites, brief structural comparisons among members, and different modes of auto-inhibition and activation.
Keywords: RING-between-RING ubiquitin E3 ligases; protein ubiquitylation; ubiquitin.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Figures
References
-
- Marin I, Lucas JI, Gradilla AC, Ferrus A. Parkin and relatives: the RBR family of ubiquitin ligases. Physiol Genomics. 2004;17:253–63. - PubMed
-
- Eiyama A, Okamoto K. PINK1/Parkin-mediated mitophagy in mammalian cells. Curr Opin Cell Biol. 2015;33:95–101. - PubMed
-
- Kitada T, Asakawa S, Hattori N, Matsumine H, Yamamura Y, Minoshima S, et al. Mutations in the parkin gene cause autosomal recessive juvenile parkinsonism. Nature. 1998;392:605–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
