

Improving the Immunogenicity of Native-like HIV-1 Envelope Trimers by Hyperstabilization
- PMID: 28834745
- PMCID: PMC5590011
- DOI: 10.1016/j.celrep.2017.07.077
Improving the Immunogenicity of Native-like HIV-1 Envelope Trimers by Hyperstabilization
Abstract
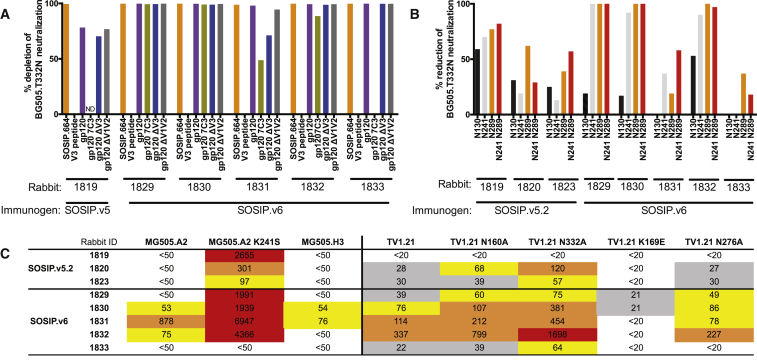
The production of native-like recombinant versions of the HIV-1 envelope glycoprotein (Env) trimer requires overcoming the natural flexibility and instability of the complex. The engineered BG505 SOSIP.664 trimer mimics the structure and antigenicity of native Env. Here, we describe how the introduction of new disulfide bonds between the glycoprotein (gp)120 and gp41 subunits of SOSIP trimers of the BG505 and other genotypes improves their stability and antigenicity, reduces their conformational flexibility, and helps maintain them in the unliganded conformation. The resulting next-generation SOSIP.v5 trimers induce strong autologous tier-2 neutralizing antibody (NAb) responses in rabbits. In addition, the BG505 SOSIP.v6 trimers induced weak heterologous NAb responses against a subset of tier-2 viruses that were not elicited by the prototype BG505 SOSIP.664. These stabilization methods can be applied to trimers from multiple genotypes as components of multivalent vaccines aimed at inducing broadly NAbs (bNAbs).
Copyright © 2017 The Authors. Published by Elsevier Inc. All rights reserved.
Figures



References
-
- Binley J.M., Sanders R.W., Clas B., Schuelke N., Master A., Guo Y., Kajumo F., Anselma D.J., Maddon P.J., Olson W.C. A recombinant human immunodeficiency virus type 1 envelope glycoprotein complex stabilized by an intermolecular disulfide bond between the gp120 and gp41 subunits is an antigenic mimic of the trimeric virion-associated structure. J. Virol. 2000;74:627–643. - PMC - PubMed
-
- Bulleid N.J., van Lith M. Redox regulation in the endoplasmic reticulum. Biochem. Soc. Trans. 2014;42:905–908. - PubMed
-
- Camacho C.J., Thirumalai D. Modeling the role of disulfide bonds in protein-folding: entropic barriers and pathways. Proteins. 1995;22:27–40. - PubMed
-
- Creighton T.E. Disulphide bonds and protein stability. BioEssays. 1988;8:57–63. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
